О ВОЗМОЖНЫХ МЕХАНИЗМАХ ФОРМИРОВАНИЯ ВОЗБУЖДАЮЩИХ И ТОРМОЗЯЩИХ КОРТИКОФУГАЛЬНЫХ ВЛИЯНИИ В ПРОЕКЦИОННОЙ КОРЕ
При рассмотрении механизмов избирательного кортикофугального контроля мы постулировали наличие возбуждающих и тормозящих влияний вторых сенсорных областей коры на входные системы передаточных ретикулярных нейронов. Обратимся к фактам, которые позволяют полагать, что такие эффекты действительно могут формироваться в проекционной коре.
Наиболее удачным доказательством было бы прямое наблюдение одновременных облегчающих и тормозящих влияний, формируемых в нейронных системах локального пункта II сомато-сенсорной области коры. Однако до сих пор нет достаточно точных признаков, по которым можно было бы судить о том, оказывает ли данная группа нейронов облегчающий или тормозящий эффект на другие структуры. Исключение составляют тормозящие интернейроны, работающие в системе возвратного торможения, реакции которых можно различать с большей уверенностью. Однако тормозящие интернейроны работают в системе «своих» нейронов, от которых они получают аксонную коллатераль; вероятность работы таких тормозящих клеток с нейронными системами других структур мозга пока еще никем не рассматривалась. Поэтому критерии, применяемые для идентификации интернейронов возвратного торможения, мы не могли использовать. Не имея возможности определить по ответным реакциям корковых нейронов характер их вероятного кортикофугального влияния, мы ограничились косвенным наблюдением (Р. А. Дуринян, 1968, 1969). Погружая одновременно два микроэлектрода в верхние (III—IV) и нижние (V—VI) слои коры II сомато-сенсорной области исследовали фоновые реакции одиночных нейронов до, во время и после охлаждения этого участка. На рис. 25 видно, что до охлаждения нейроны слоя III—IV коры имели достаточно высокий уровень фоновой активности (1—3). При охлаждении поверхности коры активность нейронов постепенно снижается (4—7) и практически исчезает полностью (8—10), когда температура коры достигает 20°С. При отогревании этого участка коры фоновая активность вновь восстанавливается (11—13) и в конце опыта приближается к первоначальной (14).
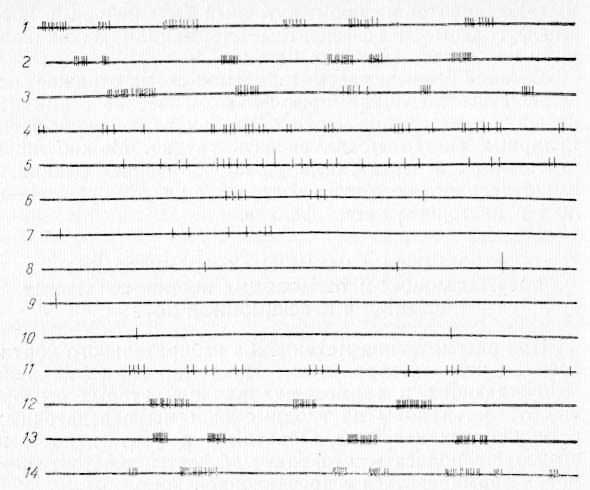
Рис. 25. Изменение фоновой активности одиночного нейрона слоя III—IV коры в области CII при ее охлаждении. 1—3 — до охлаждения; 4—10 — при охлаждении; 11—14 — при отогревании.
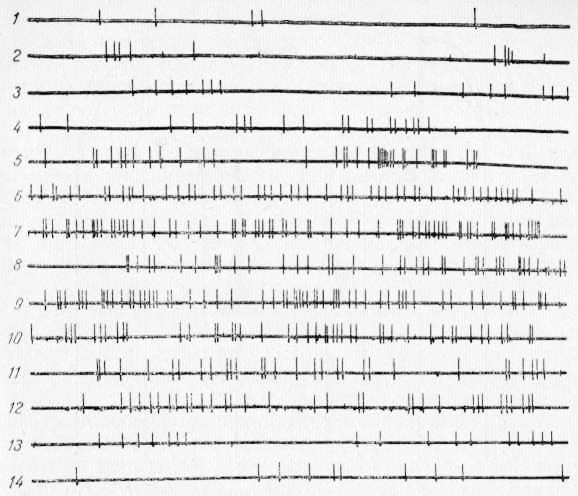
Рис. 26. Изменение фоновой активности одиночного нейрона слоя V—VI коры в области CII при ее охлаждении. 1—3 — до охлаждения; 4—10 — при охлаждении; 11—14 при отогревании.
Эту закономерность наблюдали многие авторы (Т. В. Козырева, 1972; Gartside, Lippold, 1967; Barker, Carpenter, 1970). Более интересным и важным представляется нам динамика изменения фоновой активности нейронов нижних слоев коры (рис. 26). До охлаждения (1—3) фоновая активность нейронов слоя V—VI коры достаточно низкая. При охлаждении поверхности коры резко усиливается фоновая активность этих нейронов (4—7), несмотря на то что температура в глубоких слоях коры близка к нормальной. При некотором понижении температуры (до 32°С) фоновая активность нейронов глубоких слоев (8—10) не только не снижается, но может даже усиливаться. При отогревании поверхности коры (11—13) активность этих нейронов постепенно снижается и к концу процедуры (14) возвращается к исходному уровню. Эти исследования показали, что при охлаждении поверхности коры фоновая активность нейронов нижних слоев коры резко усиливается. Объяснить это явление изменением температуры в нижних слоях нет никаких оснований, так как в начале охлаждения, когда температура там не ниже 36°С, фоновая активность уже значительно усиливается (5—7). Продолжающееся усиление фоновой активности при дальнейшем охлаждении можно отнести за счет возбуждающего эффекта некоторого понижения температуры в глубине коры к концу периода охлаждения (10). Однако при отогревании высокий уровень активности сохраняется (11—12) до тех пор, пока не восстанавливается фоновая активность нейронов верхних слоев коры. Таким образом, наблюдается явная взаимозависимость в характере фоновой активности нейронов верхних и нижних слоев коры. Фоновая активность нейронов нижних слоев коры в обычных условиях ниже, чем верхних. Это различие можно объяснить тем, что афферентная импульсация по таламокорковым проекциям поступает непосредственно к нейронам слоя III—IV. Однако это не объясняет, почему при подавлении активности нейронов верхних слоев усиливается активность нейронов нижних слоев коры. Не является ли это показателем того, что нейроны верхних слоев коры оказывают на нижележащие нейроны тормозящий эффект? К такому же предположению пришла Т. В. Козырева (1972). При подавлении активности нейронов слоя III—IV коры интенсивность их тормозящего влияния на нижележащие нейроны снижается и в связи с этим усиливается фоновая активность этих нейронов.