Ультра структур а ША и других постсинаптических специализаций подробно изучалась (А. А. Манина, Р. П. Кучеренко, 1971; Н. С. Косицын, 1973; П. К. Анохин, 1975; А. А. Манина, 1976; Eccles, 1973). В аксодендрити- ческих синапсах на шипиках ША наблюдается только у высших животных и далеко не во всех шиповидных выростах. Особенностью аксошипиковых синапсов с ША у разных животных является их исключительная пластичность и динамичность (рис. 9, см. вкл.). Цитоплазма шипикового выроста более специализирована по сравнению с дендритом, что особенно отчетливо обнаруживается при изучении ультраструктуры синалтосом (рис. 10, см. вкл.). В синаптосомах шипик часто остается прикрепленным к постсинаптической области, демонстрируя тем самым прочные адгезивные свойства синаптического контакта.
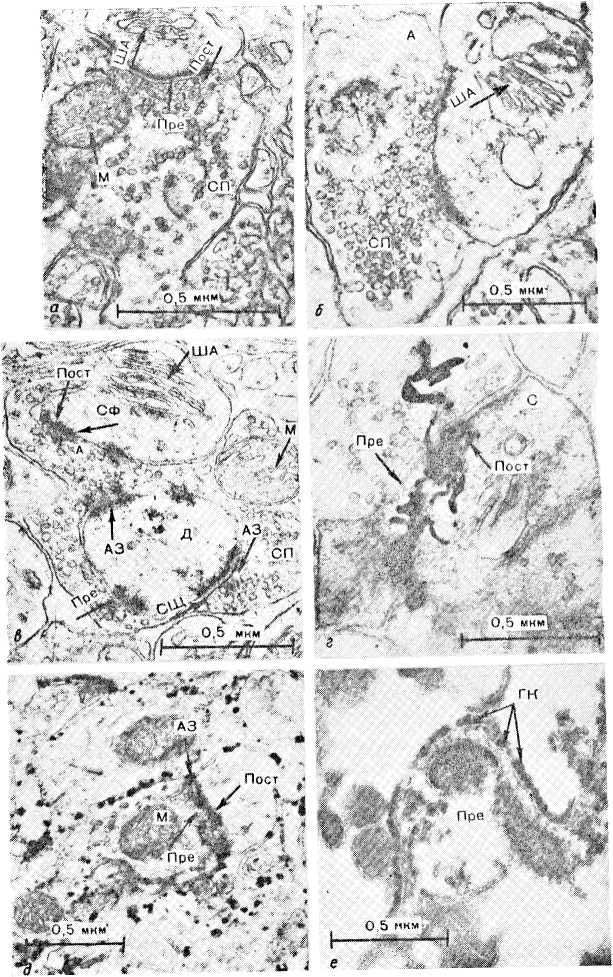
Рис. 9. Ультраструктура (а, б, в), гистохимия (д, е) и авторадиография (г) (включение 3H-лейцина) аксошипиковых синапсов, а — у обезьяны, б — у собак, в—е — у крыс. Обозначения то же, что на рис. 6; ГК — гликокаликс.
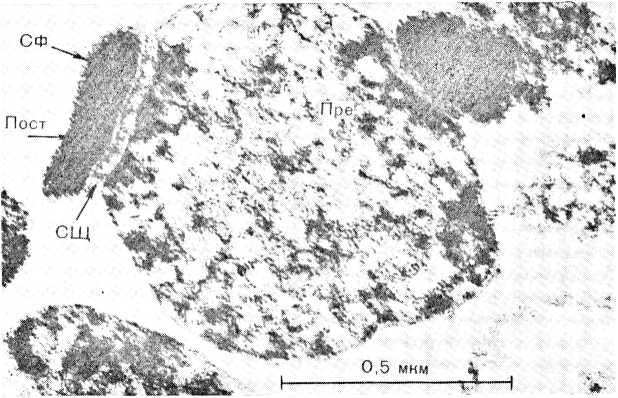
Рис. 10. Синаптосома с двумя активными зонами синапса (выделена ультра- центрифугированием). Обозначения те же, что на рис. 6.
В аксошипиковых синапсах, как и в других синапсах, в коре головного мозга отдельная синаптическая бляшка может образовывать несколько активных зон (рис. 11, см. вкл.). Такая структура осуществляет более надежную передачу нервного импульса и делает возможным варианты выбора для передачи информации и пространственно-временного распространения нервного импульса. Активные зоны синапса могут передавать сигнал одновременно или последовательно, что видно на электроно- грамме и схеме (рис. 12) (возбужденное и тормозное состояние активных зон на одном синапсе). Таким образом, даже в пределах одного синапса интеграция передачи информации осуществляется различно, что обеспечивает надежность ее проведения.
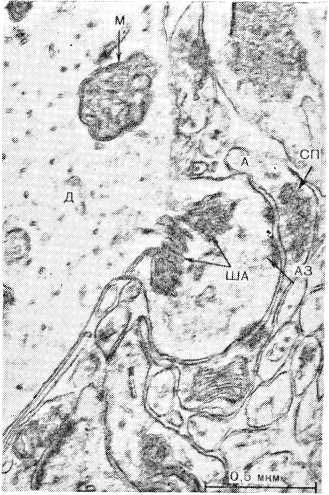
Рис. 11. Аксошипнковын синапс с ША. Генез ША от плазмолеммы дендритной терминалы. Обозначения те же, что на рис. 6.
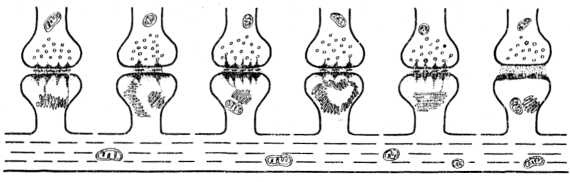
Рис. 12. Схема, иллюстрирующая расположение ША, его контакты с субсинаптической мембраной, филаментами при различном функциональном состоянии (последний слева в неактивном состоянии).
При обучении и стимуляции ЦНС фенамином в процессе образования временных связей как формы проявления памяти у различных животных повышается структурно-функциональная активность аксошипиковых синапсов. Отмечается конформационная перестройка синаптического комплекса. В постсинаптическом полюсе синапса, в шипиках, у разных животных развивается закономерная специфическая реакция — пролиферация сложных мембранных комплексов, составляющих ША. На рис. 12 представлена схема, иллюстрирующая развитие ША при мотивации. Выявлена выраженная специализация ША, который образует различное количество взаимосвязанных канальцев, окруженных мембранами и субмембранными электронно-плотными филаментами, достигающими величины 30—50 нм. Количество мембран может достигать 30 и более, а их длина варьирует от 0,25 до 1 мкм.