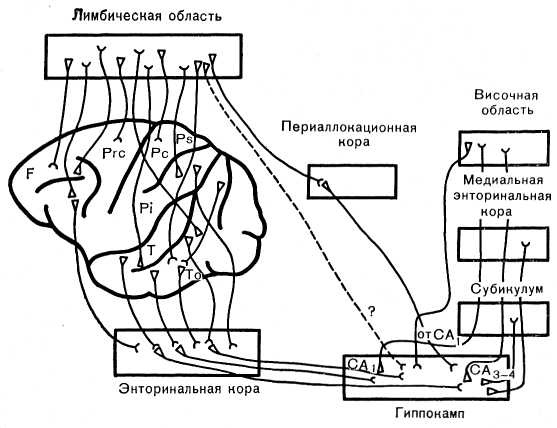
Рис. 29. Принципиальная схема ассоциативных связей дорсального неокортекса с лимбической областью коры, энторинальной корой и гиппокампом. Пунктиром обозначен путь, не подтверждаемый исследованиями последних лет.
1. Подобно межкортикальным системам, связывающим различные территории дорсального неокортекса между собой, связи последнего с лимбической и энторинальной областями коры прогрессивно развиваются в сравнительном ряду млекопитающих.
Одно из важных проявлений прогресса взаимоотношений конвекситального неокортекса с лимбической и энторинальной областями у приматов по сравнению с субприматами— увеличение удельного веса систем волокон, идущих к этим областям от вторичных проекционных и ассоциативных кортикальных территорий — лобной, височной и теменной (Showers, 1959; Nauta, 1972). Таким образом, для организации этих связей у приматов также может быть справедливо правило Flechsig (рис. 29).
2. Для связей структур конвекситального и лимбического неокортекса (как и для путей между долями новой коры на наружной поверхности полушария) характерно сочетание принципа топографического (рострокаудального) распределения ассоциативных волокон с принципом более или менее значительного их перекрытия. У субприматов орокаудальная организация ассоциативных связей с лимбической корой характерна в первую очередь для структур, находящихся над поясной извилиной на внутренней поверхности полушарий. Однако это, по-видимому, не исключается и для отдельных образований дорсолатеральной коры. У приматов топографическая организация связей дорсолатерального неокортекса с лимбической и энториальной областями еще более очерчена. Она также характеризуется зонами перекрытия (И. А. Замбржицкий, 1972; Showers, 1959; Jones, 1969).
3. Для понимания путей и механизмов межцентральной интеграции существенное значение имеет факт установления многочисленных и разнообразных ассоциативных эфферентов лимбического неокортекса. Эти эфференты направляются главным образом к неокортикальным полям наружной поверхности полушарий. По-видимому, у приматов эти связи, подобно ассоциативным афферентам лимбической коры, нарастают и для них также характерна топическая организация проекций наряду с их перекрытием.
Таким образом, в филогенезе по мере совершенствования и дифференциации неокортикальной пластики вычленения ее так называемых вторичных (проекционноассоциативных) и третичных (ассоциативных) зон создаются условия для более дробного и сложного обмена информацией за счет не только вертикальных, но также и горизонтальных систем связей. Этот вывод справедлив как для областей конвекситального неокортекса, так и для взаимоотношений его с лимбической областью коры. Дифференциация последней, как хорошо известно,также усложняется от низших млекопитающих к высшим.
4. Многие исследователи отмечают, что лимбические неокортикальные поля богаты проекционными эфферентами (рис. 30). Они идут не только к передним таламическим ядрам, посылающим в свою очередь основную афферентацию к лимбической коре, но также к другим ядрам зрительного бугра. Более того, лимбический неокортекс имеет отчетливые связи со многими образованиями ствола — ядрами моста, черной субстанцией, центральным серым веществом, субталамическими ядрами, с оградой и также с базальными ганглиями — хвостатым ядром, бледным шаром (И. А. Замбржицкий, 1972; Т. С. Сотниченко, 1973; Showera, 1959). Имеются данные о связи лимбической коры со спинным мозгом в виде лимбико-спинальных дорсального и бокового пучков (И. А. Замбржицкий, 1972). Есть указания на связь передних лимбических полей, с латеральным, гипоталамусом.
Все это позволяет утверждать, что лимбическая область коры, помимо хорошо развитых ассоциативных связей, имеет отчетливые выходы к промежуточным эффекторным и, возможно, к исполнительным аппаратам мозгового ствола и спинного мозга. Данные физиологических и клинических исследований хорошо с этим согласуются (Н. Н. Веллер, 1971; Showers, 1959). Таким образом, по нашим представлениям лимбическая область коры, особенно ее передние отделы, близка к так называемым интегративно-пусковым структурам неокортекса — моторной и орбитальной территориям коры, в которых осуществляются окончательный синтез кортикальных импульсаций и формирование основного эфферентного интеграла (О. С. Адрианов, 1970, 1976).
