Полученные факты свидетельствуют о том, что в РФ среднего мозга, помимо большой группы нейронов, активируемых через восходящие спинальные системы, имеются также нейроны, активность которых запускается по кортикофугальным системам. Среди этих «кортикофугальных» нейронов РФ имеются две разновидности. Одна группа активируется исключительно по кортикофугальным системам II сомато-сенсорной области коры. Для другой группы характерна конвергенция двух афферентных потоков: восходящих— спинальных и нисходящих — кортикофугальных. Если в отношении этих конвергентных нейронов можно говорить о тормозящих или облегчающих влияниях сомато-сенсорной коры, особенно СII, то для группы «чистых» кортикофугальных нейронов следует говорить о кортикофугальном способе их активации. В первом случае речь может идти о кортикофугальной модуляции активности ретикулярных нейронов на периферические соматические стимулы. Во втором случае речь идет о корти кофугальном способе передачи афферентных сигналов на ретикулярные нейроны. Не исключена возможность, что обе группы этих ретикулярных нейронов, которые так или иначе получают сигналы по кортико-ретикулярным связям, составляют различные звенья одной и той же системы ретикулярных нейронных ансамблей, которые участвуют в интеграции соматической информации. Возможно, в эту систему входят и те нейроны, которые отвечают только на периферические стимулы. Их возможные взаимоотношения рассмотрены в следующей главе.
При стимуляции передаточного реле таламуса (VP) одиночные нейроны РФ среднего мозга отвечают характерной реакцией, латентный период которой на 7—10 мс короче, чем при стимуляции периферических нервов (В. Л. Цатуров, Р. А. Дуринян, 1970). Например, при стимуляции нервов передней конечности ответ одиночного нейрона наступает через 19—22 мс, а при стимуляции фокуса максимальной активности зоны представительства передней конечности в VP — через 12—13 мс. При охлаждении II сомато-сенсорной области реакции одиночных нейронов РФ, вызванные стимуляцией VP, полностью прекращаются (рис. 16) и восстанавливаются при отогревании коры. При прямой стимуляции коры (Б. Т. Чувин, 1971) в области СII реакция одиночных нейронов наступает с латентным периодом 8—10 мс, т. е. на 4—5 мс быстрее, чем при стимуляции таламического реле. Эти несколько миллисекунд разницы указывают на то, что сигнал, поступивший из VP в СII коры, не направляется сразу в РФ или в СМ, а задерживается в коре. За это время, по-видимому, происходит обработка сигналов в корковых нейронах, после чего лишь формируется кортикофугальный импульс на нейронные системы РФ и СМ. Латентный период кортикофугальных реакций при регистрации микроэлектродом экстраклеточной импульсной активности одиночных нейронов на несколько миллисекунд больше, чем при отведении фокальных медленных потенциалов обычным электродом. Здесь нет никакого противоречия. Это объясняется тем, что медленный фокальный потенциал, отражающий в суммированном виде ВПСП реагирующих нейронов, развивается сразу же, с началом постсинаптической деполяризации нейронов, а разряды этих нейронов появляются лишь после достижения критического уровня деполяризации. При небольшой крутизне постсинаптической деполяризации на это уходит несколько миллисекунд, которые и составляют разницу в превышении латентного периода импульсной реакции по отношению к фокальному потенциалу этих нейронов. При большой крутизне деполяризации разность латентного периода между фокальным и импульсным ответом будет минимальной, что наблюдается на примере реакции таламических релейных нейронов при синхронном ответе большой группы.
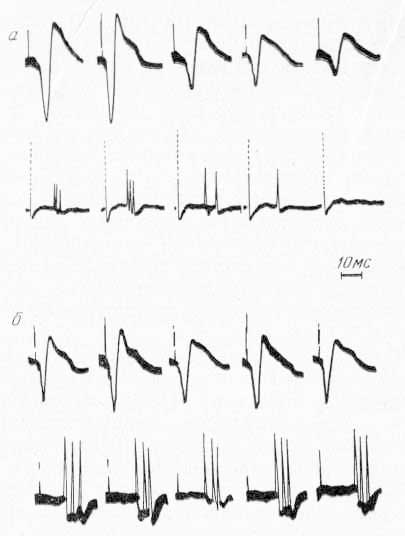
Рис. 16. Изменение экстраклеточной активности (нижний луч) одиночного нейрона ретикулярной формации среднего мозга, вызванного стимуляцией VP таламуса, при охлаждении сомато-сенсорных областей коры. Верхний луч — фокальные потенциалы в области СII коры. а — при охлаждении СII; б — при охлаждении CI.