Такая же зависимость существует в СМ и для периферических сигналов и свидетельствует о тесных функциональных взаимоотношениях обеих полушарий уже на таламическом уровне. Прямым доказательством того, что при стимуляции таламического реле наиболее вероятный путь передачи сигналов в РФ и СМ проходит через проекционную сомато-сенсорную кору, являются опыты с экстирпацией или охлаждением этой области коры. В этих условиях потенциалы исчезали полностью (рис. 7).
Афферентные проекции таламического реле, как известно, достигают обеих сомато-сенсорных областей, однако выявить, какая из этих областей ответственна за передачу возбуждения в исследуемые структуры, методом экстирпации невозможно. Даже локальное повреждение одной из соматических областей коры (отсасывание или термокоагуляция) является настолько травмирующей процедурой, что неизменно приводит к угнетению первичного ответа в другой области (контроль при стимуляции нервов и таламического ядра). Тем не менее даже такой грубый прием показал, что связь через СII более тесная.
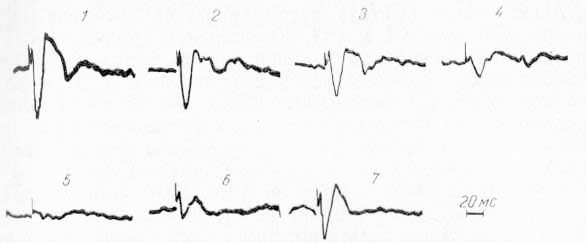
Рис. 7. Потенциалы, отводимые от СМ таламуса, при стимуляции VP таламуса в норме и при охлаждении области СII коры. 1 — до охлаждения; 2—5 — во время охлаждения; 6—7 — при отогревании.
Некоторые представления о преимуществах кортико- фугальных связей области СII с РФ среднего мозга и СМ таламуса получены косвенным путем, при исследовании фокальных потенциалов этих структур, вызванных стимуляцией периферических нервов. Тогда уже было отмечено, что в отдельных случаях наблюдается прямая зависимость амплитуды ретикулярных ответов от амплитуды первичных ответов в фокусе максимальной активности II сомато-сенсорной области коры. В отношении CI такая зависимость не выявлялась. Это полностью совпадает с той закономерностью, которая имела место при стимуляции таламического реле. Принимая во внимание это обстоятельство, мы решили получить более убедительные доказательства, подтверждающие преимущества кортикофугальной системы II сомато-сенсорной области.
Наиболее удачной моделью, достаточно близкой к естественным условиям, оказалась именно стимуляция заднего вентрального ядра таламуса. Дело в том, что разные участки этого ядра по-разному связаны с сомато-сенсорными областями коры. Заднее вентральное ядро (VP) состоит из двух половин: VPL и VPM. Латеральная часть (VPL) всеми без исключения признается как реле для I сомато-сенсорной области. Meдиальная часть (VPM) рассматривается как реле одновременно для CI и СII. Каудальный полюс VPM и так называемые ядра задней группы служат реле только для II сомато-сенсорной области. Несмотря на то что вопрос о передаточных ядрах для CI и СII полностью нельзя считать решенным, в большинстве работ последних лет именно так рассматривается классификация таламических реле сомато-сенсорной коры (Аndersson, 1962; Mehler, 1965; De Vito, 1967; Jones, 1967; Jones, Powell, 1968; Rinvik, 1968). Достаточно четко были идентифицированы проекции этих ядер в кору при помощи их локальной стимуляции. Топическое распределение ответов наблюдалось не только в CI, но и в СII (Guillery, Adrian, Woolsey, Rose, 1966); было обнаружено большое число нейронов, посылающих проекции одновременно к обеим областям (Andersen, Аndersson, Landgren, 1966; Andersson, Landgren, Wolsk, 1966).