Падение амплитуды коркового ответа при охлаждении, как это показал А. И. Ройтбак (1964), сопровождается постепенным увеличением длительности отдельных его фаз. На этом фоне хорошо проявляются высокочастотные колебания на положительной фазе, отражающие разряд таламо-кортикальных терминален (см. рис. 10,4—6). Эти колебания сохраняются и после исчезновения первичного ответа и свидетельствуют о том, что афферентные сигналы продолжают поступать в кору. Падение ответа в одной области при охлаждении другой объясняется постепенным охлаждением и другой области й связано, по-видимому, с понижением температуры циркулирующей крови. Поскольку кровоток направлен от СII к CI (Kawamura, 1971), падение потенциалов CI при охлаждении СII наступает значительно быстрее, чем в области СII при охлаждении CI. Более быстрое восстановление ответов в СII после ее предварительного охлаждения можно, вероятно, объяснить лучшими условиями кровоснабжения и большей устойчивостью ее нейронов к различным физическим и химическим воздействиям (О. Г. Баклаваджян, Ф. А. Адамян, 1964).
Для нас было важно убедиться, что при локальном охлаждении функциональное выключение обеих областей коры происходит неодновременно, что при охлаждении CI подавление ответов в СII наступает значительно позже того, когда ответы в CI уже полностью подавлены. Эти факты позволили, подавляя попеременно активность сомато-сенсорных областей коры, проследить зависимость ретикулярных ответов от величины потенциалов в CI или СII.
Основные эксперименты этой серии (Р. А. Дуринян, А. Г. Рабин, 1963а, б; А. Г. Рабин, Р. А. Дуринян, 1963) проводились в следующей последовательности. Стимулирующий (биополярный, концентрический) электрод погружали в заднее вентральное ядро таламуса в зону представительства передней конечности. Регистрируя потенциалы в CI и СII, находили в ядре такой участок, стимуляция которого вызывала максимальные первичные ответы в обеих областях коры. Затем погружали отводящие электроды в РФ среднего мозга и в СМ таламуса. Регистрирующая аппаратура, к сожалению, не позволяла одновременно отводить потенциалы от 4 точек мозга. Мы записывали их попеременно. Например, при охлаждении СII потенциалы отводили сначала в CI и в СМ таламуса; после отогревания, повторяя процедуру охлаждения, отводили потенциалы в CI и в РФ среднего мозга; время от времени проверяли потенциалы в СII.
При локальном охлаждении II сомато-сенсорной области коры, обычно на 2-й минуте, происходит постепенное падение амплитуды потенциалов срединного центра и ретикулярной формации среднего мозга, вызванных стимуляцией заднего вентрального ядра таламуса.
К этому моменту первичный ответ в I сомато-сенсорной области не только не уменьшается, но даже несколько увеличивается (рис. 11,2—4), а в области СII ответы уже резко подавлены. Спустя еще 1 — Н/г мин, при продолжающемся охлаждении СII потенциалы исследуемых структур исчезают полностью, хотя в области CI первичный ответ еще достаточно выражен (рис. 11,5—6).
В области СII ответы уже не отводятся. Наконец, падает амплитуда ответа в CI и сохраняется лишь небольшое положительное колебание (рис. 11,7), отражающее приход в кору афферентного залпа. При отогревании чаще всего ответ в CI быстро восстанавливается, в то время как в СМ и даже РФ это происходит медленнее (см. рис. 11,5—10).
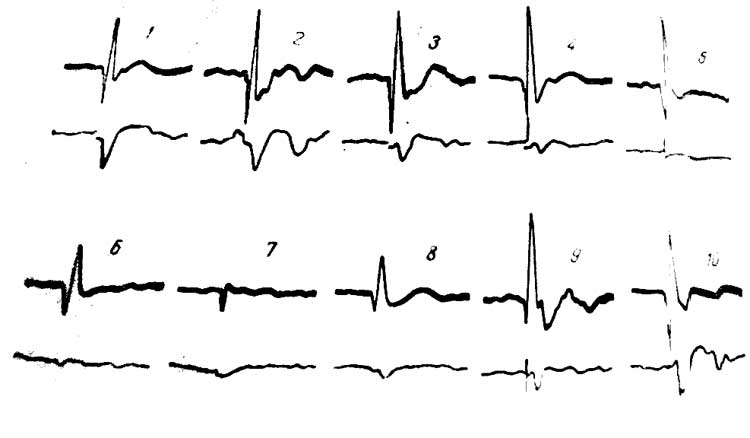
Рис. 11. Изменение потенциалов СМ (нижний луч), вызванных стимуляцией VP таламуса при охлаждении СII области коры. Верхний луч — запись потенциалов CI. 1 — до охлаждения; 2—7 — во время охлаждения; 8—10 — при отогревании.
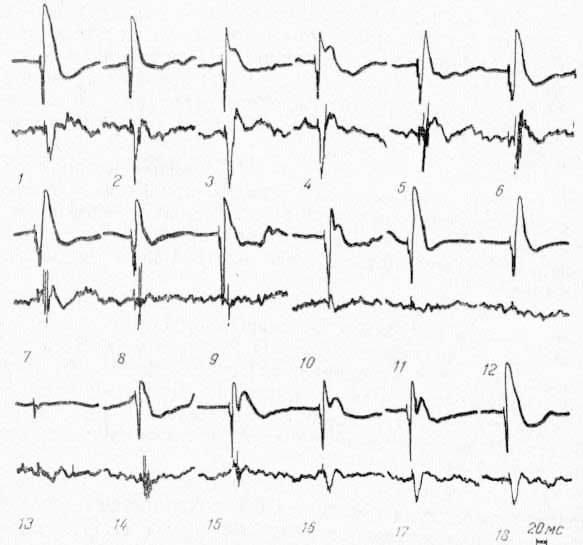
Рис. 12. Изменение потенциалов ретикулярной формации среднего мозга (нижний луч), вызванных стимуляцией VP таламуса, при охлаждении CII области коры. Верхний луч — запись потенциалов CI. 1 — до охлаждения; 2 — 13 — во время охлаждения; 14 — 18 — при отогревании.
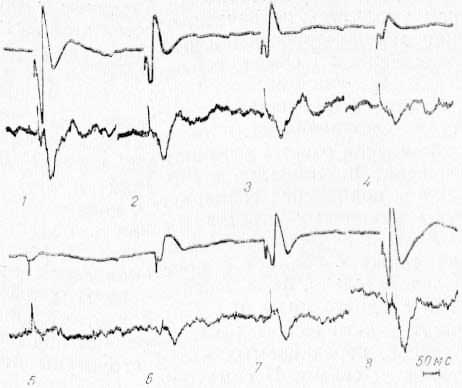
Рис. 13. Изменение потенциалов ретикулярном формации среднего мозга (нижний луч) вызванных стимуляцией VP таламуса, при охлаждении CI области коры. Верхний луч — запись потенциалов СII. 1 — до охлаждения; 2-5 — во время охлаждения; 6—8 — при отогревании.
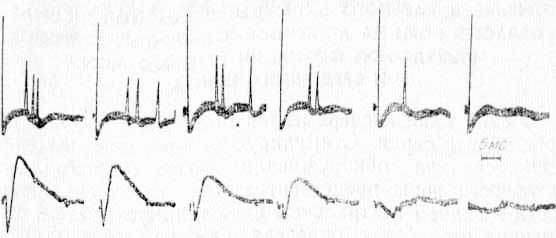
Рис. 14. Изменение экстраклеточной активности (верхний луч) одиночного нейрона СМ таламуса, вызванного стимуляцией VP таламуса, при охлаждении СII области коры. Нижний луч — фокальные потенциалы в области охлаждения СII.
Динамика изменения потенциалов в РФ представлена на рис. 12. Эти эксперименты показали, что между амплитудой потенциалов СМ и РФ и амплитудой первичных ответов CI нет явной зависимости. Падение амплитуды потенциалов этих структур н их восстановление происходят параллельно с падением и восстановлением амплитуды первичных ответов в области СII. Указанная закономерность наблюдалась также в опытах с охлаждением I сомато-сенсорной области. В этом случае, как уже указывалось, падение амплитуды первичных ответов в СII начинается спустя 8—10 мин после начала охлаждения CI, в то время как в самой области охлаждения ответы исчезают уже через 2—3 мин.
Уменьшение потенциалов в РФ (как и СМ) и их почти полное подавление совпадают с началом падения амплитуды первичных ответов именно в области СII (рис. 13,2—5), а не охлаждаемой области CI; восстановление их также совпадает с восстановлением ответов в СII. Следовательно, фокальные потенциалы, вызываемые в РФ среднего мозга и в СМ при стимуляции передаточного таламического реле, возникают под влиянием импульсов, передаваемых в эти структуры по кортикофугальным связям II сомато-сенсорной области коры. Этот вывод подтвердился также при исследовании экстраклеточной активности одиночных нейронов СМ и РФ.