Ионные насосы известны не только для натрия и калия, но и для других веществ. Способность этих насосов «перекачивать» ионы против градиента концентрации просто фантастична. Так, оказалось, что изолированная кожа лягушки перекачивает поваренную соль из раствора, омывающего ее наружную поверхность, в раствор, соприкасающийся с внутренней поверхностью, даже тогда, когда концентрация ее во внутреннем растворе в 10 000 раз больше, чем в наружном. Еще пример — через 1 см2 мембраны нерва кальмара перекачивается 10 млрд, ионов натрия в секунду. На эту работу затрачивается около 20% всей энергии, производимой клеткой. Коэффициент же полезного действия таких насосов достигает 50—95%. Такого к. п. д. не имеет ни одна самая совершенная машина, созданная человеком.
Есть предположение, что существует не общий переносчик ионов натрия и калия, а для каждого свой собственный или один, но способный превращаться то в переносчик ионов калия, то натрия (рис. 34).
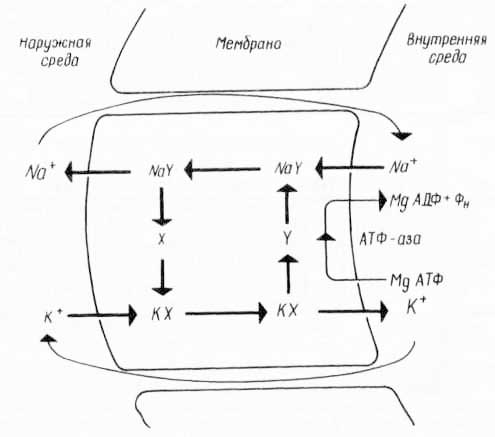
Рис. 34. Предполагаемый механизм «калий-натриевого насоса» с двумя переносчиками X и Y (из К. Вилли, В. Детье, 1974).
Предполагается также, что активный перенос ионов через биологические мембраны осуществляется за счет биохимических процессов и благодаря особенностям расположения в мембране биохимических ансамблей белков, ферментов и др. Примером может быть дыхательный ансамбль мембраны митохондрий. Эта мощная «машина» способна не только к образованию АТФ, но и к перемещению ионов к конформационным изменениям мембран. Последние, вероятно, связаны и с конформационными изменениями углеводородных цепей фосфолипидов, и с движением белков в мембранах (рис. 35). Существуют данные, что все перечисленные активности являются «векторными». Для этого должна быть соответствующая структурная основа, т. е. асимметричная организация индивидуальных ферментов или целых ансамблей в мембране. Если мембрана сама по себе асимметрична (к примеру, одна из ее поверхностей более гидрофильна), то перенос молекул будет носить векторный характер. На рис. 36 изображена модель мембраны, где фиксируются молекулы фермента с асимметричным активным центром. Дыхательные ансамбли также характеризуются асимметричной организацией (рис. 37).
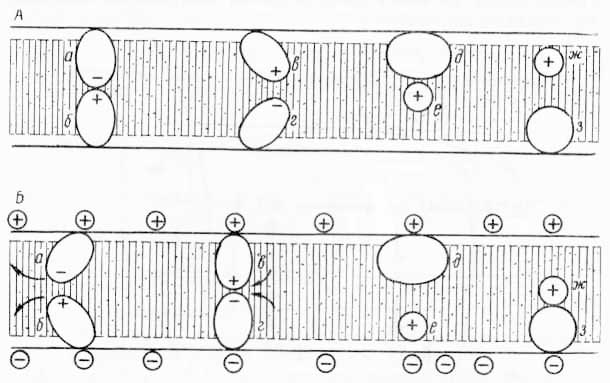
Рис. 35. Предполагаемая схема воздействия электрического поля на контакт «жестких» белковых глобул в мембране: А — наиболее устойчивое постулируемое состояние мембранных белков в отсутствие трансмембранного поля; Б — состояние белков при появлении электрического поля; а, б — глобулы, расходящиеся (обозначено стрелками) иод действием электростатического притяжения разноименных зарядов; в, г — глобулы, приходящие в контакт в результате отталкивания одноименных зарядов белков и поверхностей мембраны; д, е — глобулы, разъединяющиеся (ж, з — соединяющиеся) из-за миграции одной из них (е и ж).

Рис. 36. Модель векторной реакции (из А. Ленинджера, 1974): верхняя схема — асимметричное образование ионов Н+ и ОН— в процессе переноса электронов; средняя схема — мембранная АТФ-аза, способная к асимметричному образованию (связыванию) Н+ и ОН— (или фосфолипидных групп); нижняя схема — модификация, объясняющая асимметричность образования ионов Н+ и ОН— и АТФ в каждом пункте сопряжения.
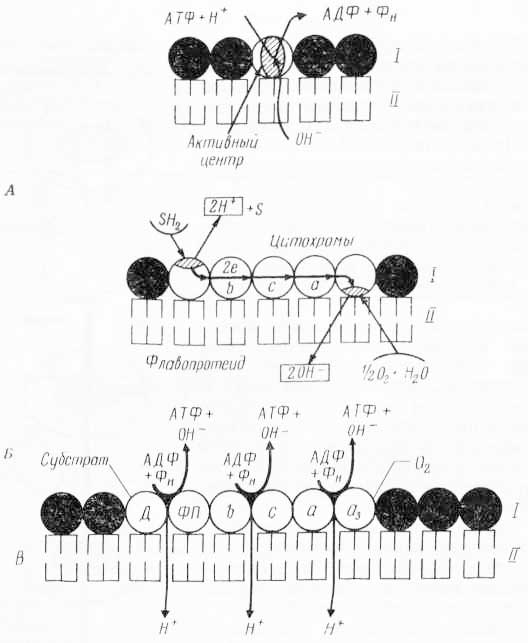
Рас. 37. Асимметричное расположение активных центров АТФ-аз и переносчиков электронов Н+ и ОН— в мембранной структуре относительно непроницаемой для ионов Н+ (схема механизма переноса ионов) (по А. Ленинджеру, 1966): А — асимметричная АТФ-аза, создающая градиент pH; Б — разделение ионов Н+ и ОН— в процессе окисления Н2-субстрата кислородом, обусловленное асимметричным и противоположным расположением флавопротеида и цитохромоксидазы в мембране; В — АТФ-аза, ответственная за окислительное фосфорилирование АДФ с одновременным разделением ионов Н+ и ОН—; I — белковый слой, II — липидный слой.
Таким образом, действительно существуют механизмы («насосы») активного переноса через мембрану не только ионов калия и натрия, но и ионов водорода и гидроксильной группы, т. е. кислоты.
ОБЩИЕ ПРИНЦИПЫ РЕАЛИЗАЦИИ ВЗАИМООТНОШЕНИЙ В СИСТЕМЕ ОРГАНИЗМ — СРЕДА
Главная » Энциклопедия » Физиология » Книги по физиологии » Книги по общей физиологии » Главы книг по общей физиологии » ОБЩИЕ ПРИНЦИПЫ РЕАЛИЗАЦИИ ВЗАИМООТНОШЕНИЙ В СИСТЕМЕ ОРГАНИЗМ — СРЕДА
Страницы: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40