Мы полагаем, что изменение или модуляция пропускной способности главного (передаточного) полимодального ретикулярного нейрона происходит еще до поступления афферентных периферических сигналов на этот главный нейрон и осуществляется на группе вставочных нейронов. Это те нейроны, которые отвечают на стимуляцию коры и периферических афферентов. На рис. 23 представлена схема предполагаемых взаимоотношений между группой ретикулярных нейронов, которая с минимальным противоречием объясняет механизм реализации избирательного коркового контроля на афферентную вызванную активность главных ретикулярных нейронов.
На этой схеме рассматриваются два типа нейронов. Первый (более крупный) является главным ретикулярным нейроном, на который конвергируют сигналы разных сенсорных модальностей и который передает информацию в другие структуры мозга, в частности в кору больших полушарий. Это своего рода передаточный нейрон (n) ретикулярной формации, который в отличие от передаточных нейронов специфических ядер-реле способен потенциально передавать сигналы разной модальности. В определенных случаях под влиянием коры этот нейрон «специализируется» на передаче сигналов какой-то одной сенсорной модальности.
Нейроны меньшего размера рассматриваются как «входные» и промежуточные элементы для главного (передаточного) ретикулярного нейрона. Эти клетки активируются только сигналами одной и той же сенсорной модальности: либо по периферическим афферентам (а) с разной протяженностью рецептивного поля, либо по кортикофугальным путям (к), либо двойным способом — афферентным и кортикофугальным (ак). На схеме указан один главный нейрон, хотя вполне возможно, что промежуточные нейроны объединяют в один комплекс группу таких главных (передаточных) ретикулярных нейронов. На гистологических препаратах, окрашенных по методу Гольджи (3. В. Елисеева, 1969), можно видеть множество таких комплексов в СМ таламуса, где несколько более крупных нейронов с длинными дендритами окружены группой мелких клеток с менее длинными дендритами. Аксоны этих мелких клеток не уходят за пределы поля зрения и сосредоточены вблизи крупных клеток (рис. 24).

Рис. 23. Схема взаимоотношений главного передаточного (п) ретикулярного нейрона с соответствующими «входными» (в) интернейронами, объясняющая механизм избирательного коркового контроля. А — без выделения отдельных типов интернейронов; Сомат — соматические интернейроны; Зрит.— зрительный интернейрон; Слух.— слуховой интернейрон; ипс. — ипсилатеральный; контр. — контралатеральный; а — афферентные тер- минали соответствующих систем; о — облегчающие и m — тормозящие терминали кортикофугальных проекций из соответствующих одноименных корковых областей (соматической, зрительной, слуховой); Б — с выделением трех типов интернейронов входа для каждой проекционной системы (на примере соматической системы): к — кортикофугальный интернейрон с облегчающей (о) и тормозящей (т) кортикофугальными терминаля- ми; ак — смешанный афферентно-кортикофугальный интернейрон с возможными тормозящими (т) кортикофугальными терминалями; « — афферентный интернейрон с соответствующим периферическим афферентным входом. Обозначены также окончания других (Зрит., Слух, и т. п.) смешанных ак интернейронов на передаточном (n) ретикулярном нейроне.
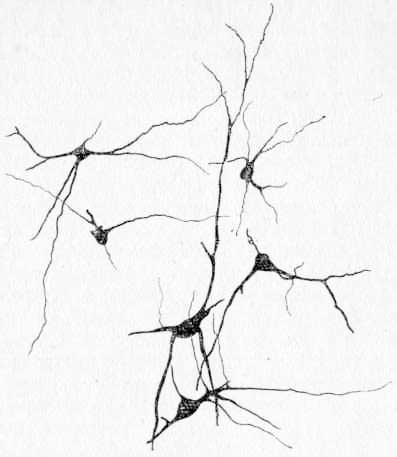
Рис. 24. Пример взаимоотношения нейронов СМ таламуса кошки (зарисовка с препарата, окрашенного по Гольджи).
Рассматриваемая схема состоит из двух последовательных вариантов. Первый, упрощенный, вариант (см. рис. 23, А) отражает общий механизм избирательного коркового контроля на главный ретикулярный нейрон без учета существования отдельных «входных» корти- кофугальных и афферентных нейронов.
На рис. 23, Б дан второй более сложный вариант с учетом трех типов промежуточных нейронов у каждого входа. Рассмотрим схему более подробно. На рис. 23, А указан один ретикулярный передаточный нейрон (n) и 4 промежуточных, или вставочных, нейрона (в). Аксоны вставочных нейронов образуют синапсы на передаточном нейроне, по-видимому, аксо-дендритного типа. Вставочные нейроны, указанные вверху рисунка, являются «входными» системами для соматических (Сомат.) сигналов: слева ипсилатеральных (ипс.); справа контралатеральных (Контр.). Два других вставочных нейрона, расположенные справа ниже, представляют собой входные системы для зрительного (Зрит.) и слухового (Слух.) сигналов. Ипсилатеральные входы для них не указаны.
На каждом вставочном нейроне представлены следующие синаптические системы: кортикофугальные облегчающие (о), по-видимому, аксо-дендритного типа, кортикофугальные тормозящие (т), по-видимому, аксо-соматического типа и восходящие афферентные (а) аксо-дендритного типа.