Вызванные потенциалы ядра R таламуса. Трудности, связанные с точным попаданием кончика электрода в это ядро, осложняли электрофизиологическое исследование.
В литературе имеются лишь отдельные указания па возможность отведения соматических потенциалов в этом ядре (Kruger, Albe-Fessard, 1960).
В связи с этим мы особое внимание обратили на изучение характеристики вызванных потенциалов в ядре R таламуса. В этих экспериментах (1966) регулярно проводился гистологический контроль расположения кончика отводящего электрода в ядре R. Потенциалы ядра R, подобно потенциалам других ретикулярных структур, представляют собой двухфазное колебание. Начально положительная волна ответа достигает амплитуды 300 мкВ и переходит в большую отрицательную волну порядка 150 — 200 мкВ. Перед положительной фазой часто виден небольшой отрицательный пик, как и у потенциалов других структур, который, как уже указывалось, отражает активность соседнего специфического реле таламуса. Латентный период ответов на 2 — 3 мс больше, чем в ретикулярной формации среднего мозга, и близок к латентному периоду ответов комплекса СМ — Pf на аналогичные стимулы. Например, при стимуляции нервов передней конечности латентный период в ядре R равен 12 — 14 мс. В отличие от потенциалов СМ — Pf и особенно ретикулярной формации положительная фаза ответа ядра R имеет меньшую длительность, которая не превышает 20 мс. При медленной развертке луча осциллографа может возникнуть впечатление о сходстве этого ответа с потенциалами специфического реле. Однако при большой развертке легко установить, что временные параметры этого потенциала превышают соответствующие показатели первичного ответа заднего вентрального ядра. Так, например, при стимуляции нервов передней конечности латентный период ответа VP равен 5 — 6 мс, а длительность положительной фазы 8 — 12 мс. При раздражении ипсилатеральных нервов латентный период ответов в ядре R на 2 — 3 мс больше, чем контралатеральных стимулов, а амплитуда ответов значительно меньше.
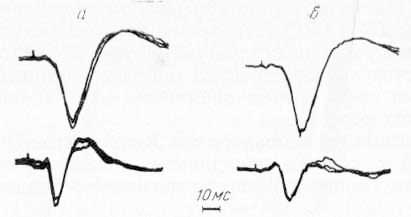
Рис. 3. Потенциалы ретикулярной формации среднего мозга (верхний луч) и ретикулярного ядра таламуса (нижний луч), вызванные стимуляцией контралатеральных (а) и ипсилатеральных (б) соматических нервов.
Как видно на рис. 3, для ретикулярной формации среднего мозга практически нет различий между контра- и ипсилатеральными ответами. В таламическом отделе эти различия становятся достаточно заметными, что свидетельствует о более концентрированной проекции контралатеральных афферентных терминалей на нейронных элементах СМ — Pf.
Хотя «латерализация» соматических афферентных проекций в ядрах неспецифического таламуса менее строгая, чем в специфическом передаточном ядре лем- нисковой системы, тем не менее она свидетельствует о более выраженной специализации афферентных систем в таламических неспецифических ядрах по сравнению с ретикулярной формацией. Об этом же свидетельствует меньшая длительность положительной фазы ответа в неспецифических ядрах таламуса, особенно заметная для ядра R. Уже на основании этих данных можно думать о более компактных и локализованных проекциях соматических афферентных систем в комплексе СМ — Pf и в ядре R таламуса, по сравнению с ретикулярной формацией среднего мозга, что вполне согласуется с морфологическими данными. Относительно локальные зоны представительства афферентных систем в неспецифическом таламусе вполне могут являться основанием для более локальных проекций в кору и потенциальными источниками для менее диффузных, концентрированных реакций в коре при адекватных раздражениях рецепторных систем организма.