Наши эксперименты показали, что у крыс, находящихся в возбужденном состоянии под действием фенамина, синаптосомальный комплекс претерпевает изменения, отражающие конформационную перестройку синаптических мембран, связанную с усилением медиации и трансмембранным транспортом ионов. Использование ФВК и нитрата лантана позволило локализовать углеводные биополимеры на поверхности нейрональных мембран.
Высказано предположение, что в межнейрональных контактах коры головного мозга важную роль играют аксодендритические синапсы с шипиками дендритов (Г. Н. Поляков, 1953; С. А. Саркисов, Н. Н. Боголепов, 1967; Cajal, 1935; Eecles, 1973). В механизмах формирования энграмм памяти Szentagothai (1971), Eccles (1973) придают исключительное значение шипикам. Они предполагают, что продолжительность электрической активности в постсинаптической области в процессе обучения связана со структурными изменениями в дендритных шипиках и что высокий уровень электрической активности — следствие гипертрофии (рис. 8), почкования или деления шипиков. Эти исследователи считают, что следовые процессы фиксируются на шипиках в виде сложных множественных энграмм при обучении, являющихся копиями паттернов сигнала, который используется для считывания и извлечения информации. Однако эти авторы признают, что для подтверждения их гипотезы необходимы электронно-микроскопические доказательства. Несомненно, как представляют эти исследователи, гипертрофия или деление шипиков в процессе обучения могли бы привести лишь к развитию патологии мозга.
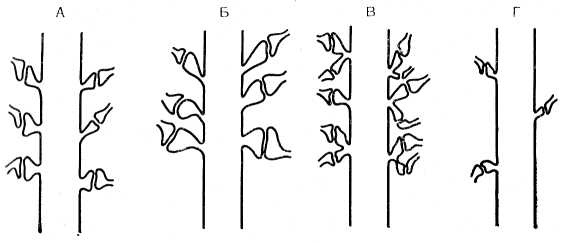
Рис. 8. Схема, иллюстрирующая реакцию ША при формировании энграмм памяти (Eccles, 1973).
А — норма шипиков; Б — гипертрофия шипиков; В — деление шипиков; Г — утрата функции шипиков.