Другой системой, претендующей на роль длинного ассоциативного пути, является поясной пучок (cingulum). Он располагается у основания сводчатой извилины над мозолистым телом. Большинство исследователей считают его смешанным: длинные волокна его образуют проекционную систему центрального обонятельного нейрона у собаки (Г. Я. Трошин, 1903) и человека (С. Б. Дзугаева, 1975). Удаляя в свое время моторную область коры у собак, мы (О. С. Адрианов, 1952) обнаружили дегенерации в поясном пучке, идущие довольно далеко назад, однако их нельзя было заметить за пределами этого пучка. Наша сотрудница Е. С. Неклюдова (1965), используя метод расщипывания нервных волокон, показала, что у обезьян (макак резусов) поясной пучок истончается по направлению к передним отделам полушария, а его компактная часть огибает валик мозолистого тела (рис. 27). Pfuhl (1955) рассматривает поясной пучок в качестве ассоциационного пути, связывающего область орбитально-поясной коры с гиппокампом, а также в качестве пути, соединяющего филогенетически старейшие части конечного мозга — миндалину и гиппо- камповую формацию с филогенетически новейшими областями лобной коры. Ряд авторов (Т. С. Сотниченко, 1968; White, 1959; Domesick, 1969), используя современные импрегнационные методики, пришли к заключению о проекции поясного пучка не на структуры гиппокампа, а на энторинальную и пресубикулярную кору.
Короткие ассоциативные волокна, входящие в состав поясного пучка, связывают различные участки лимбической области коры в переднезаднем направлении (П. И. Яковлев, 1967; И. А. Замбржицкий, 1972; С. Б. Дзугаева, 1975). Полагают, что в составе поясного пучка проходят афферентные пути от таламических ядер к лимбическому неокортексу, пресубикулум и энтори- нальной коре (Shipley, 1974). П. И. Яковлев (1967), И. А. Замбржицкий (1972), Т. С. Сотниченко (1973), Showers (1959) считают, что через поясной пучок проходит большое число ассоциативных эфферентов лимбической коры к различным областям наружной поверхности полушария.
Большинство исследований отражают наличие у субприматов длинных ассоциативных систем связей, которые соединяли бы передние и задние отделы неокортекса. По данным А. Л. Микеладзе и Г. И. Кикнадзе (1972), длинные ассоциативные системы волокон, связывающие лобную область с теменной, появляются у собак, но еще отсутствуют у кошек. Kawamura (1973а, b, с), Kawamuга и Otani (1970) также не смогли найти прямые связи между лобной областью неокортекса кошки, с одной стороны, и слуховой, зрительной и теменной зонами — с другой.
В свое время мы (О. С. Адрианов, 1952), используя методику Марки, нашли, что ассоциативные связи моторной области коры мозга собаки не распространялись за пределы соматосенсорных полей. Позднее нами (О. С. Адрианов, 1970, 1976) был проведен детальный морфофизиологический анализ взаимодействия передних и задних половин неокортекса при образовании у собак различных по характеру условных рефлексов на простые и сложные раздражители. Морфологически исследовался мозг подопытных собак, подвергшихся операции полного двустороннего разобщения передних половин полушария от задних (через всю толщу коры и подлежащего белого вещества полушарий мозга вплоть до бокового желудочка). При окраске мозга на волокна по методике Шпильмейера у всех без исключения животных было отмечено серьезное побледнение волокон белого вещества полушарий по обе стороны от уровня разреза, но не более чем на 8—12 мм кпереди и кзади от линии разобщения. Более тонкий анализ возможности существования ассоциативных связей между передними и задними половинами полушарий был проведен нами с помощью методов Марки и Наута. Эти данные в принципе подтвердили наши прежние результаты. Перерожденные волокна шли от разреза компактным путем, а потом постепенно рассеивались.
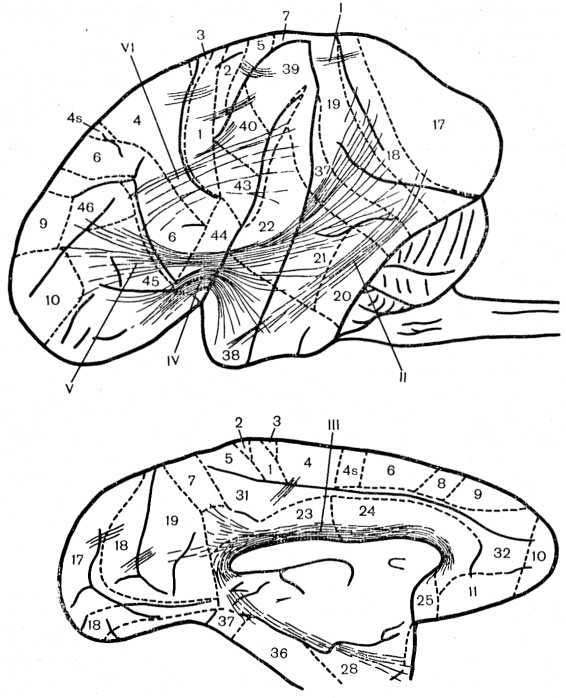
Рис. 27. Схема длинных ассоциативных путей полушарий мозга обезьяны макаки резуса (по Е. С. Неклюдовой, 1966).
I—III — обозначения те же, что на рис. 26; IV — крючковидный пучок; V — лобно-затылочный пучок; VI — дугообразный пучок.