ВЛИЯНИЕ ЛОКАЛЬНОГО ОХЛАЖДЕНИЯ СОМАТО-СЕНСОРНЫХ ОБЛАСТЕЙ КОРЫ НА АКТИВНОСТЬ ОДИНОЧНЫХ НЕЙРОНОВ РЕТИКУЛЯРНОЙ ФОРМАЦИИ СРЕДНЕГО МОЗГА И СРЕДИННОГО ЦЕНТРА
Общий план экспериментов был таким же, как и в предыдущей серии. Стимулируя нервы плечевого сплетения, сначала обнаруживали фокус максимальной активности зоны представительства передней конечности в заднем вентральном ядре таламуса; затем, стимулируя этот фокус, определяли фокус максимальной активности зоны представительства в I и II сомато-сенсорных областях коры. После этого вводили микроэлектрод в СМ или РФ, приступали к попеременному охлаждению CI или СII и исследовали экстраклеточ- ную активность одиночных нейронов этих структур.
При охлаждении СII (Р. А. Дуринян, 1968) экстраклеточная активность нейронов СМ, вызванная стимуляцией ядра VP, полностью прекращалась. После отогревания коры вызванная активность этих нейронов вновь восстанавливалась (рис. 14). То же самое происходило при стимуляции нервов.
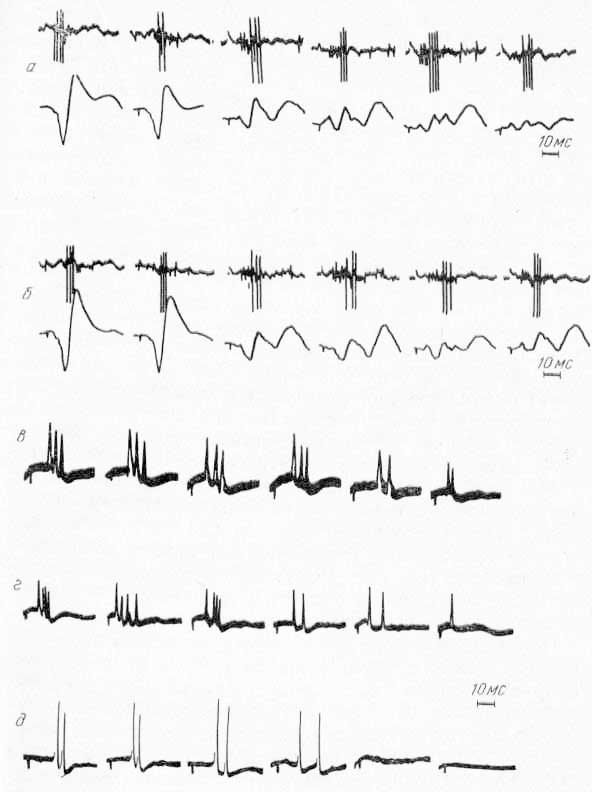
Рис. 15. Реакция одиночных нейронов ретикулярной формации среднего мозга, вызванных стимуляцией нервов, при охлаждении сомато-сенсорных областей коры. а, б — сохранение реакции нейронов РФ (верхняя запись) при охлаждении CI (а) и СII (б). Нижняя запись — фокальные потенциалы в точках охлаждения Коры; в г — частичное изменение реакции нейронов РФ при охлаждении CI (в) и особенно СII (г); д — подавление реакции нейронов РФ при охлаждении СII.
В ретикулярной формации среднего мозга наблюдалась несколько иная картина (В. Л. Цатуров, Р. А. Дуринян, 1968). Реакции примерно 50% исследованных ретикулярных нейронов, вызываемые стимуляцией нервов, не изменялись сколько-нибудь заметно при охлаждении CI и СII. Даже аппликация 2% раствора хлористого калия оказалась неэффективной. Нейроны такого типа, по-видимому, активируются только афферентными импульсами по спино-таламическим и спино-ретикулярным путям (рис. 15, а, б). У другой группы нейронов охлаждение CI и СП приводило к уменьшению числа спайков в ответе нейрона, но полностью ответ не исчезал. Особенно заметно подавлялась реакция нейронов этого типа при охлаждении СII на стимуляцию контралатеральных нервов. Стимуляция ипсалатеральных нервов продолжала вызывать обычную реакцию (рис. 15, в, г). После отогревания коры исходная реакция нейронов восстанавливалась. Этот факт свидетельствует о том, что при стимуляции соматических нервов активация этой группы ретикулярных нейронов происходит частично по кортико-ретикулярным путям, помимо активации по афферентным спинальным путям.
Наконец, третья группа нейронов РФ среднего мозга отвечает исключительно только на кортикофугальный разряд. При охлаждении области СII активность этих нейронов постепенно снижается, а затем полностью прекращается (см. рис. 15, 6). Восстановление происходит после отогревания области СII, и тогда эти нейроны начинают вновь отвечать на стимуляцию нервов.