Однако столь необходимый для жизнедеятельности ритм при старении и возрастной патологии выходит из строя. Действительно, как можно видеть на рис. 25, построенном на основании полученных в нашей лаборатории данных, стандартная доза глюкозы вызывает (в соответствии со своей ролью в энергетическом гомеостазе) падение уровня гормона роста в крови у молодых лиц, но не дает этого важнейшего эффекта ни у мужчин среднего возраста, ни при такой характерной возрастной патологии, как рак молочной железы и рак тела матки у женщин. Это означает, что гипоталамус проявляет резистентность к тормозящему гомеостатическому действию глюкозы. Именно наличие резистентности создает нарушение ритма в смене источников энергии при поступлении в организм углеводов. Можно видеть, что в ответ на введение глюкозы не только не происходит снижения продукции гормона роста у лиц с возрастными изменениями, но не снижается и концентрация в крови жирных кислот. Иными словами, несмотря на поступление пищи извне и увеличение в крови концентрации глюкозы, концентрация жирных кислот тоже остается высокой, т. е. происходит нарушение ритма в работе энергетического гомеостата».
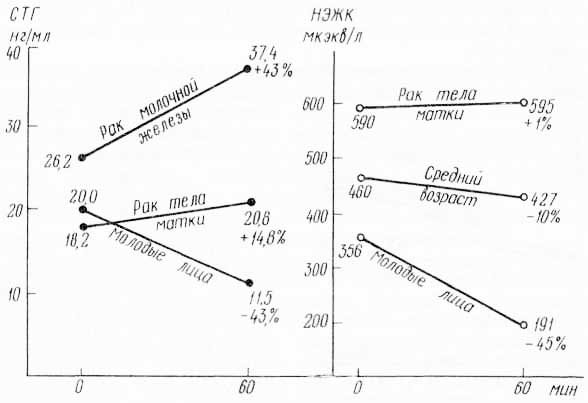
Рис. 25. Эндокринный гомеостаз (по В. М. Дильману, 1972).
Попытки привести все сказанное к каким-то количественным оценкам предпринимались неоднократно. Но цель настоящих очерков не сказать все, а попробовать сказать обо всем в самом обобщенном, принципиальном виде.[9] Поэтому следует познакомиться с одной из наиболее общих моделей процесса конечного развития по Прохоренко (рис. 26).
1. Кумулятивный обмен обеспечивает накопление субстрата, измеренного по какому-то параметру z, в системе по логистическому закону. На участке молодого экспоненциального роста, простирающегося от t=0 до t=t1 будем иметь:
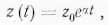
где z0≠0 и означает величину параметра z при t = 0, когда начинается экспоненциальный рост; α — постоянная экспоненциального роста.
2. Мощность внешнего обмена u(t)в соответствии с основным свойством кумулятивности определяется количеством наличного субстрата в системе и в каждый момент времени пропорциональна ему:
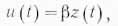
где β — удельная мощность внешнего обмена.
3. Поставляемый внешним обменом субстрат реализуется внутренним обменом по двум существенно различным каналам: истинный обмен (обновление наличного субстрата) с мощностью u0(t), также определяемой количеством наличного субстрата и пропорциональной последнему:
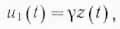
где Υ — постоянная удельная мощность истинного обмена и, во-вторых, обмен роста (кумулятивное накопление субстрата) системы с мощностью u2(t), равной скорости роста параметра z и выражающейся через производную функцию z по времени:
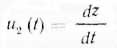
В стадии экспоненциального роста последнее уравнение дает:
![]()
и постоянная α приобретает смысл удельной мощности роста. Записываем баланс мощностей обмена:
![]()
который должен выполняться по крайней мере в период раннего развития. Подставляя значения мощностей u1, u2, u3,получим:
![]()
Кривые мощности обмена строим следующим образом. Допустим β=1, а α=Υ, тогда кривая внешнего обмена u воспроизводит график z. Дифференцируя графически логистическую кривую u, находим мощность роста u1. Кривую мощности истинного обмена u1 строим, удовлетворяя баланс мощностей обмена на участке экспоненциального роста. Суммируя кривые u1 и u2, получим мощность полного внутреннего обмена u1 + u2 как функцию времени. Графическим дифференцированием кривой u2 находим ускорение роста системы u2.
[9] Автор приносит свои извинения за чрезмерную самонадеянность.