Исключительного развития в клетках достигла система мембран. Клеточная мембрана не только отграничивает одну клетку от другой, но и образует многочисленные впячивания в цитоплазму: от очень просто устроенных (пиноцитозные вакуоли-пузырьки) до сложнейшей эндоплазматической сети. Полагают, что эндоплазматическая сеть выполняет весьма важную функцию в осуществлении обмена веществ между цитоплазмой и окружающей клетку средой.
Таким образом, клетка состоит из двух не смешивающихся друг с другом фаз: из основного вещества цитоплазмы со всеми возможными в ней переходами от жидкого до твердого состояния и из мембранной системы, расчленяющей цитоплазму на своеобразные отсеки, получившие название цитоплазматического матрикса.
Следовательно, не только клетки пространственно отделены друг от друга, но и их структуры (рис. 28). Разрушение мембран приводит к уничтожению клетки как самостоятельной единицы.
Значительную часть вещества, из которого формируется основная структура биологических мембран, составляют фосфолипиды. Модель молекулы фосфолипида изображена на рис. 29. Как видно из рисунка, молекула фосфолипидов состоит из гидрофильной головки и двух гидрофобных «хвостов» (цепей углеводородов), что создает ее асимметричность как по химической структуре и пространственному расположению частей, так и по избирательному отношению к тем средам (фазам), с которыми она соприкасается. К поверхности воды молекулы фосфолипидов ориентируются гидрофильными головками, в то время как гидрофобные «хвосты» будут обращены в обратную сторону от нее. В этом легко убедиться на примере поведения молекул полярных липидов (рис. 30, 31, 32). Известно, что элементарная биологическая мембрана состоит из двух слоев фосфолипидов и их гидрофильные головки обращены к периферии, поскольку снаружи мембрану той или иной клетки окружает межклеточная жидкость — внешняя водная фаза, а внутренняя — цитоплазма также представляет собой водную фазу. В аналогичных условиях находятся и все органеллы клеток. Следовательно, биологические мембраны отделяют друг от друга две различные по составу и свойствам водные фазы.
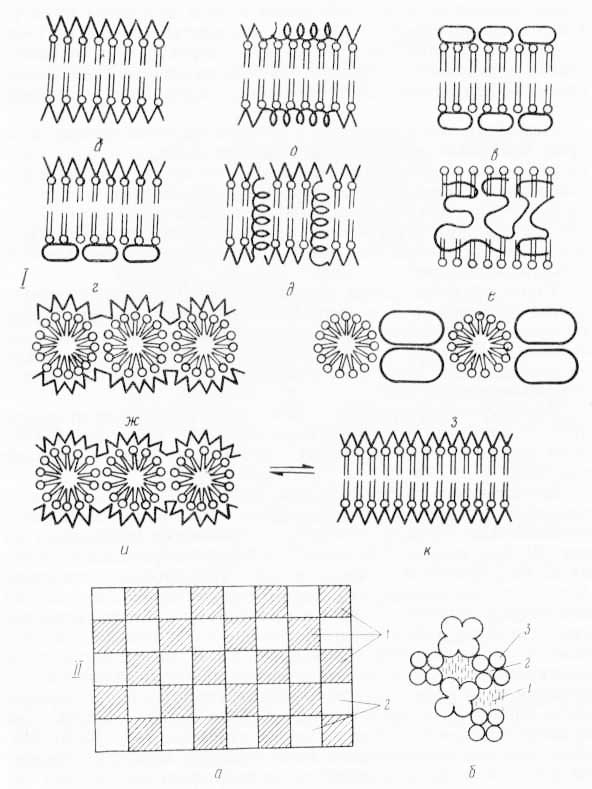
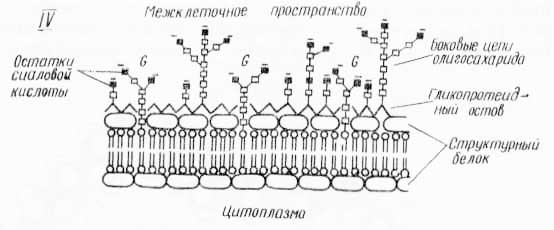
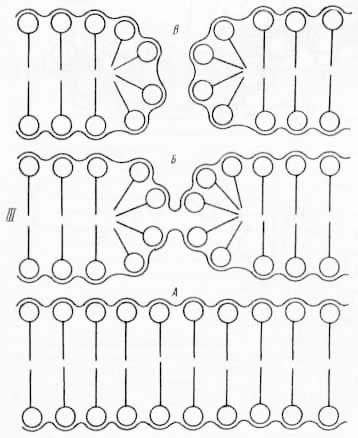
Рис. 28. Различные модели молекулярного строения биологических мембран: I. В основу моделей положены: двухслойная липидная структура (а — е); глобулярная организация (ж — к); α — белок в β-форме; б — α-спираль; в — глобулярный белок; г — асимметрия в расположении белка; д — канальцы и поры, частично пронизывающие белковый слой; е — белок внутри двойного слоя липидов; ж — липидные мицеллы с 3-белком; з — липидные мицеллы с глобулярным белком; и, к — превращение глобулярной организации в двухслойную. Во всех случаях фосфолипидные белковые слои поверхностей мембран не симметричны друг другу, еще более асимметрично по составу и свойствам расположение ферментных и других функциональных групп (по Э. де Робертису, В. Новинскому, Ф. Саэсу, 1973). II. Схема строения мембраны по Валлаху (из С. В. Конева и др., 1973): а — схематический вид поверхности мембраны (1 — ориентированные липиды; 2 — белки); б — продольный разрез мембраны по ее средней гидрофобной части (1 — липидный цилиндр, 2 — водный канал, 3 — белковая палочка). III. А — непрерывная мембрана; Б — начало образования прерывистой мембраны; В — конец образования прерывистой мембраны (из Л. Поликара, 1972). IV. Модель большой мембраны, включающей наружную поверхность оболочки клетки (из Э. де Робертиса, В. Новинского, Ф. Саэса, 1973).
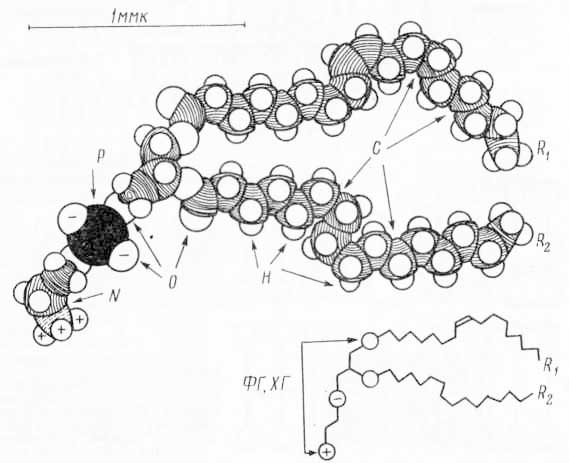
Рис. 29. Полная молекулярная модель фосфолипида (молекулы фосфатидилэтаноламина): R1, R2 — углеводородные цепи олеиновой и пальметиновой кислот; ХГ — характеристическая группа, определяющая принадлежность к данному виду фосфолипидов; ФГ — группа, соединяющая характеристическую группу с двумя углеводородными цепями; — — одинарная связь, = — двойная связь между атомами углерода; ∘ — группы, участвующие в водородной связи.
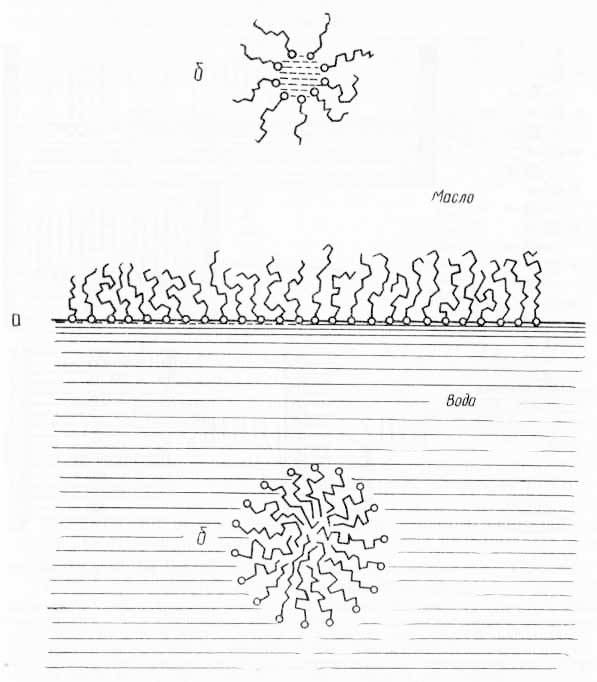
Рис. 30. Схема расположения полярных липидов (из Ж. Хаггиса, Д. Михи и др., 1967): а — расположение липидов на поверхности раздела масло — вода; б — сферические мицеллы в масляной и водной фазах.
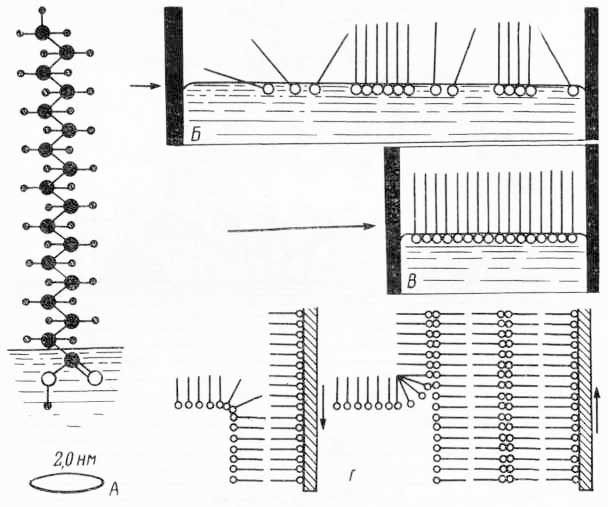
Рис. 31. Схема образования молекулярных пленок (из Э. де Робертиса, В. Новинского, Ф. Саэса, 1973): А — молекула стеариновой кислоты с полярной группой, погруженной в воду; расположение молекул: Б — при слабом сдавливании; В — при сильном сдавливании. Степень упорядоченности молекул зависит от их количества, приходящегося на площадь поверхности соприкосновения с водой и в большей мере — от выраженности их асимметрии (чем менее они симметричны, тем более упорядочено их расположение в ионослое), так как по мере увеличения последней усиливаются гидрофильные свойства отдельных частей мо лекул; Г — при погружении стеклянной пластинки с мономолекулярной пленкой стеарата бария второй молекулярный слой соединяется с первым при помощи неполярных цепей и образуется бимолекулярная пленка. Справа показано образование нескольких бимолекулярных слоев стеарата бария на стеклянной пластинке после ее многократных погружений в воду.
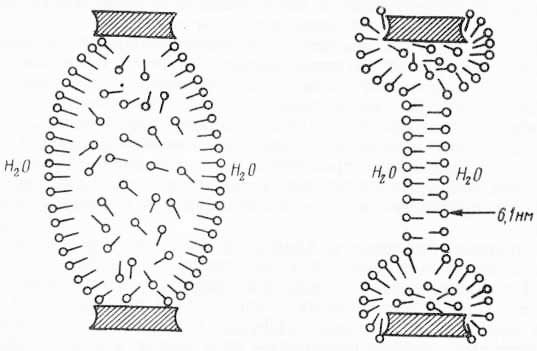
Рис. 32. Образование бимолекулярного слоя путем выталкивания воды из толстой пленки (по Томпсону и А. Поликару, 1972).
ОБЩИЕ ПРИНЦИПЫ РЕАЛИЗАЦИИ ВЗАИМООТНОШЕНИЙ В СИСТЕМЕ ОРГАНИЗМ — СРЕДА
Главная » Энциклопедия » Физиология » Книги по физиологии » Книги по общей физиологии » Главы книг по общей физиологии » ОБЩИЕ ПРИНЦИПЫ РЕАЛИЗАЦИИ ВЗАИМООТНОШЕНИЙ В СИСТЕМЕ ОРГАНИЗМ — СРЕДА
Страницы: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40