Таким образом, состояние так называемого физиологического покоя, хотя внешне и не проявляет специфической деятельности (например, движение, секреция и др.), в действительности не бездеятельно. Состояние относительно устойчивого физиологического покоя живой клетки находится в прямой зависимости от относительно устойчивого значения метаболизма и потенциала покоя. Сохранение последнего и есть один из основных показателей состояния ее физиологического покоя. Снижение потенциала покоя до нуля свидетельствует о катастрофическом нарушении проницаемости клеточной мембраны, в результате чего концентрация одноименных ионов на наружной и внутренней поверхности становится одинаковой. Возникает ионная симметрия мембраны. Это говорит о потере мембраной способности к избирательной проницаемости, о полном уравновешивании внутреннего содержимого клетки с окружающей средой, поглощении ее средой, т. е. о смерти. Вот почему потенциал покоя является одной из основных характеристик живой клетки, а следовательно, и организм в целом.
Если воспользоваться современными методами, то можно заметить, что на одном и том же участке клеточной мембраны потенциал покоя то уменьшается (гипополяризация), то увеличивается (гиперполяризация). Как известно, величина потенциала покоя зависит от разности концентрации ионов на поверхностях мембраны. Чем больше эта разность, тем больше поляризована мембрана и выше потенциал покоя. Эта разность поддерживается избирательной проницаемостью мембраны и процессами активного транспорта. В тех участках мембраны, где изменяется обмен веществ (в результате изменения состава межклеточной жидкости, вдыхаемого воздуха, температурных и других воздействий), наблюдается изменение проницаемости ее. Это сказывается на состоянии всей клетки и может быть зарегистрировано в виде колебания потенциала покоя. Так, по мере роста силы раздражения до определенного предела в месте нанесения его происходит градуально (соответственно увеличению силы раздражения) нарастающее уменьшение потенциала покоя (рис. 44). Это возможно только при уменьшении уровня поляризации поверхностей клеточной мембраны, что осуществимо лишь при условии увеличения ее проницаемости для всех потенциалобразующих ионов и в первую очередь натрия. Проникающие внутрь клетки катионы натрия уменьшают величину положительного заряда на наружной поверхности мембраны и в целом потенциала покоя. Прекращение раздражения ведет к восстановлению его исходной величины. В связи с тем что эти изменения распространяются от места своего возникновения, постепенно ослабляясь вплоть до исчезновения, а в мышечной и нервной ткани вообще ограничены небольшим участком нанесения раздражения, они и получили название местного потенциала. Важнейшим свойством местного потенциала является градуальная зависимость от силы раздражения, какой бы малой она ни была. Благодаря этому биосистема находится в состоянии постоянной готовности к реагированию на всякое воздействие. Следовательно, даже самые незначительные события небезразличны для живых организмов. Это свойство находится в полном соответствии с термодинамическими свойствами открытых стационарных и неравновесных систем.
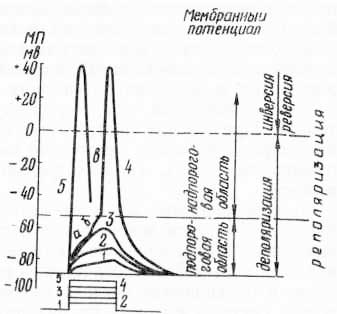
Рис. 44. Изменение мембранного потенциала в зависимости от силы наносимого раздражения длительностью 2 мс (по Б. Катцу, 1968 и Е. К. Жукову, 1969): 1—5 — изменение мембранного потенциала. Внизу — относительная сила наносимого раздражения (электрический ток). Горизонтальные линии: сплошная — уровень потенциала покоя; штрихпунктирная — критический уровень местного потенциала (подпороговая область), т. е. момент возникновения потенциала действия; штриховая—уровень перехода деполяризации в инверсию знака потенциала действия (надпороговая область); а, б — местный потенциал, включающий локальный потенциал возбуждения б; в — потенциал действия.
Другой не менее существенной особенностью местного потенциала является отсутствие латентного периода при ответе на любое воздействие, что обеспечивает максимальную оперативность в установлении наиболее оптимальных отношений организма с внешней средой и, следовательно, сохранение гомеостаза. В этом отношении замечателен тот факт, что для клеток соединительной и эпителиальной тканей, т. е. тканей, из которых в основном образованы почти все наши внутренние органы, единственной формой электрического ответа на воздействия извне являются градуальные изменения потенциала покоя, достижение критического уровня (вероятно, близкого по своей природе к явлению местного подпорогового ответа нервной и мышечной тканей) которого сопровождается возникновением выраженной функциональной активности биосистемы (для железы, например, секреция). Да как может быть по-другому, если благодаря работе именно внутренних органов обеспечивается строгое поддержание постоянства внутренней среды (состав плазмы, лимфы, межклеточной жидкости). Естественно, что ткани, их образующие, практически не могут находиться в состоянии физиологического покоя. Они должны быть постоянно в деятельности, в «готовности № 1!». Неудивительно, что местная реакция на раздражение вызвана местным возбуждением, где местный потенциал будет одним из выражений его, а величина потенциала в определенной степени — мерой возбуждения, т. е. мерой функциональной активности тканей. Причем под ростом величины возбуждения следует понимать не только и не столько внешнее его проявление (например, секреция), сколько усиление интенсивности обмена веществ.