В ходе развития местного возбуждения в отличие от распространяющегося возбуждения отсутствует состояние так называемой рефрактерности — снижения возбудимости. В этом заложен большой биологический смысл. Ведь местное возбуждение в большинстве случаев скорее характеризует готовность живой системы к ответу на раздражение, чем его непосредственное осуществление. Это относится и к тканям, для которых местный потенциал является единственной электрической характеристикой состояния возбуждения. В данном случае местный потенциал, достигнув критического уровня (возможно, сходного с локальным потенциалом возбуждения, свойственным нервной и мышечной тканям), вызывает специфическую ответную реакцию органов, состоящих из эпителиальной, железистой и соединительной тканей. Рефрактерность, свойственная одиночному циклу (волна) распространяющегося возбуждения, дает возможность полноценной реализации ответной реакции на пороговое и надпороговое раздражение. Присутствие рефрактерности в местном возбуждении так называемых возбудимых тканей (нервные клетки, мышцы) лишило бы биологическую систему способности вообще реагировать специфической ответной реакцией на раздражения.
Наконец, еще одним важным свойством местного возбуждения является ограниченность его распространения, т. е. декрементность. Говоря о местном характере возбуждения, мы имеем в виду лишь количественную сторону его, да и то в пределах возможностей измерительной аппаратуры. Теоретически нераспространяющихся и ограниченно распространяющихся событий в биологической системе нет в силу ее морфофункциональной целостности. При изучении электрических свойств мышечной и нервной тканей было замечено, что зависимость местного потенциала от силы раздражителя при достижении определенной величины нарушается (см. рис. 44). Как только местный потенциал достигает некоторой критической (пороговой) величины, он превращается в потенциал действия, что говорит о возникновении распространяющегося возбуждения.
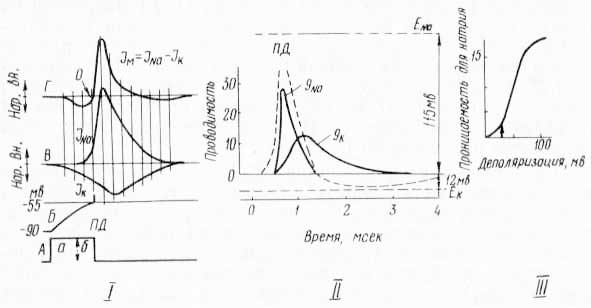
Рис. 45. Механизм возникновения потенциала действия (по Е. К. Жукову, 1969; Ходжин и Хаксли, 1952): I. А — раздражающий толчок, Б — возникновение потенциала действия; 90 мВ — уровень потенциала покоя; 55 мВ —уровень порогового (критического) потенциала; 90—55 мВ — местный потенциал: стрелка и ПД — момент возникновения потенциала действия; В — ионные потоки: нар. — направленные из клетки, вн. — внутрь; Г — суммарный ионный поток через мембрану. Вертикальные линии связывают одновременные точки на кривых ионных потоков. O — «нуль-точка» суммарного потока. II. qK, qNa — кривые изменения проницаемости мембраны для ионов калия и натрия во время развития потенциала действия; ПД — потенциал действия, мВ; ENa, EK — уровни потенциалов равновесия для ионов натрия и калия. III. Вертикальная стрелка — момент порогового сдвига проницаемости.
Потенциал действия есть следствие дальнейшего развития ионных пертурбаций на поверхностях мембраны под влиянием возросшей силы раздражения (рис. 45). Так, в подпороговой области приблизительно в течение 2/3 времени (I, А, а) от момента нанесения порогового раздражения развитие местного потенциала (I, Б) идет в условиях усиления выходящего потока ионов калия из клетки (I, В, JK) из-за снижения мембранного потенциала ЕM относительно потенциала равновесия для калия (II, ЕK). «Входящий» же поток ионов натрия в клетку (I, В, JNa) еще незначителен, так как проницаемость мембраны для них (II, qNa) пока невелика; в этих условиях суммарный поток ионов калия и натрия (I, Г) направлен из клетки наружу. По мере усиления деполяризации проницаемость мембраны для ионов натрия растет. Рост этот начинается в последней трети времени действия порогового раздражения (I, А, б); когда местный потенциал достигает критического (порогового) уровня (I, Б, ПД), встречные потоки ионов натрия и калия уравновешиваются (I, В) и суммарный поток ионов становится равным нулю (I, Г, О).
Нуль-точка суммарного потока (I, Г, О) ионов — это момент возникновения неустойчивого равновесия, т. е. неустойчивого стационарного состояния. В этих условиях даже незначительная дальнейшая деполяризация мембраны (обязательная при действии порогового раздражения) нарушает равновесие. Проницаемость мембраны для ионов натрия повышается в сотни раз (II, qNa). В результате поток их в клетку усиливается настолько, что начинает превышать поток ионов калия из клетки (I, В). Вследствие этого суммарный поток меняет свое направление— теперь он входит в клетку (I, Г) и практически не изменяется под влиянием раздражений любой сверхпороговой силы. Местный же потенциал круто преобразуется в восходящее колено потенциала действия (I, Г, Д, П, ПД). Объяснение этому можно найти, если принять предположение, что здесь регенеративный процесс идет за счет собственных зарядов мембраны клетки. Стремительно проникающий поток ионов натрия в клетку способствует и ускорению деполяризации мембраны. В результате в определенный момент мембранный потенциал становится равным нулю. Однако такое состояние (если оно практически вообще возможно) сохраняется очень короткое, не поддающееся учету время. Дело в том, что продолжающееся поступление ионов натрия в клетку не только «гасит» избыток имеющихся там отрицательно заряженных анионов, но и приводит к накоплению положительно заряженных катионов на внутренней поверхности мембраны и к недостатку их на наружной. В итоге в месте нанесения раздражения на наружной поверхности мембраны заряд меняется с положительного на отрицательный, а на внутренней — наоборот, с отрицательного на положительный, т. е. происходит изменение (инверсия) знака потенциала мембраны. При достижении потенциалом действия максимума (II, ПД), находящегося в прямой зависимости от количества ионов, перешедших с наружной поверхности мембраны на внутреннюю, поток ионов в клетку ослабляется, так как точка максимума потенциала действия близка по величине к потенциалу равновесия для ионов натрия (II, ENa). Одновременно для этих ионов начинает снижаться проницаемость мембраны (I, В, П, qNa), что способствует ослаблению суммарного потока ионов (I, Г).