При таких условиях благодаря существующему концентрационному градиенту какое-то количество ионов калия выходит из клетки через поры мембраны, создавая тем самым на ее наружной поверхности избыток положительно заряженных частиц. На внутренней же поверхности возникает избыток крупных молекул органических анионов, оставшихся без нейтрализующих их калий-положительных ионов. Благодаря электростатическим силам вышедшие катионы калия не могут далеко удалиться от наружной поверхности мембраны (положительно заряженные частицы, находящиеся вне клетки, прижимают их к мембране, а скопившиеся на внутренней поверхности отрицательно заряженные частицы стремятся «втащить» их внутрь клетки). Однако высокий концентрационный градиент для калия препятствует этому. В результате на поверхностях клеточной мембраны и возникает разность потенциалов — потенциал покоя (рис. 43). Разумеется, калий играет здесь, так сказать, основную, ведущую роль. Вместе с тем по градиенту концентрации в клетку поступает и какое-то количество катионов натрия. Но положительный заряд потока натрия значительно меньше противоположного потока ионов калия. Более того, некоторое количество анионов хлора, захватываемых натрием и поступающих вместе с органическими анионами, также участвует в образовании слоя отрицательных зарядов на внутренней поверхности мембраны, а оставшиеся в избытке катионы натрия и калия — слоя положительных зарядов на наружной поверхности мембраны. (Надо сказать, что в возникновении и поддержании потенциала покоя существенную роль играет и активный перенос, т. е. работа «калий-натриевого насоса».)
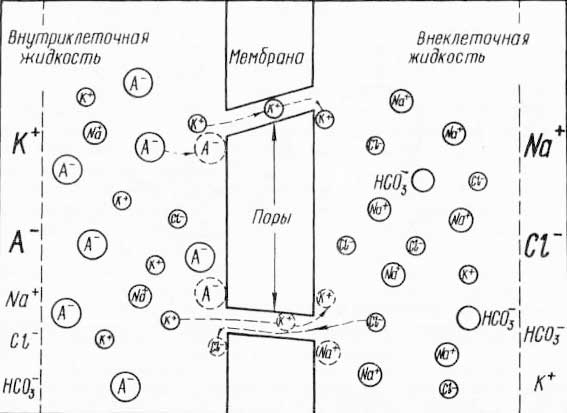
Рис. 43. Образование мембранного потенциала за счет градиента концентрации (по Вудсбери, 1962): размерами символов К+, Na+, Cl—. А- и других обозначена степень относительной концентрации соответствующих ионов.
На образование двойного электрического слоя идет очень немного ионов. Поэтому концентрационный градиент сохраняется высоким. Следовательно, диффузия ионов должна продолжаться. Однако в условиях уже существующего электрического потенциала диффундирующие ионы будут испытывать препятствие со стороны электрических зарядов на поверхности мембраны: положительные ограничивают дальнейшее поступление из клетки катионов калия, а отрицательные — приток в клетку анионов. В результате возникает состояние, при котором силы диффузии (концентрационного градиента) и электростатического отталкивания уравновешиваются: количество ионов, проходящих через мембрану по градиенту концентрации, становится равным количеству ионов, проходящих в обратном направлении под влиянием сил электростатического отталкивания. Уровень этого электрохимического равновесия и будет определять как ионную асимметрию по обе стороны мембраны (клетка — среда), так и величину мембранного потенциала.
Следует обратить особое внимание на земечательный факт возникновения и поддержания потенциала покоя как активного саморегулирующегося процесса. Вот почему потенциал покоя специфичен для различных тканей и колеблется, например, в клетках скелетных мышц в пределах 80—90 мВ, а эпителиальной ткани — 20—30 мВ. Потенциал покоя одной и той же ткани претерпевает изменения в зависимости от возраста, состояния обмена веществ и т. д., а также определяется особенностями проницаемости клеточной мембраны, изменения концентрации ионов внутри и вне клетки и др.
Проницаемость мембраны в свою очередь зависит от характера отношений в ней водных и липидных фаз. В зоне между этими фазами ионы преодолевают силы гидратации. Запас свободной энергии ионов зависит от разности энергии их при переходе из одной фазы в другую. Таким образом, при миграции частиц через биологическую мембрану возникает определенный энергетический барьер. За счет этого барьера и создается избирательная проницаемость и высокое электрическое сопротивление клеточных мембран.
Регулятором, определяющим пропускную способность мембран и разделение встречных потоков анионов и катионов, является электрический заряд поры мембраны. Привлекающая своей простотой и ясностью мембранная теория возникновения электропотенциалов клетки встречает значительные трудности при объяснении многих вопросов, связанных с выяснением их природы.