Исследования Rossi и Brodal (1956), выполненные на кошках, позволили более точно установить места окончаний кортикофугальных волокон в ретикулярной формации. Авторы отметили (рис. 1, г), что кортико-ретикулярные волокна, хотя и начинаются во многих областях коры, однако большая часть их берет свое начало в сенсомоторной коре, особенно в моторной ее части; значительно меньшее число волокон начинается в базальных и медиальных областях коры и совсем мало —- в височной и затылочной областях. Основная часть волокон идет в гомолатеральную часть ретикулярной формации, и лишь ничтожное количество их перекрещивается и направляется в контрлатеральную часть ретикулярной формации. Проекционные соматические сенсорные области входят в число тех корковых участков, которые являются источниками большого числа кортико-ретикулярных волокон (Kuypers, 1960). Это обстоятельство имеет для нас существенное значение, так как сомато-сенсорную кору мы рассматриваем как важнейший аппарат модуляции нейрональных реакций ретикулярной формации.
Как мы указывали, у всех, кто исследовал кортико-ретикулярные связи, сложилось впечатление, что большинство волокон берет начало в моторной коре. Там же начинается и пирамидный тракт. При удалении моторной коры с целью вызвать дегенерацию кортикофугальных волокон естественно, что перерождение начинает развиваться во всех нисходящих волокнах независимо от того, принадлежат они пирамидному тракту или кортико-ретикулярной системе.
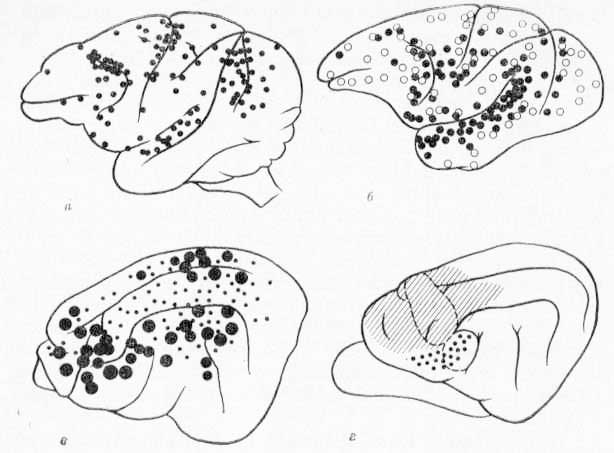
Рис. 1. Схема поверхности коры больших полушарии мозга с указанием активных кортикофугальиых участков. а — кора мозга обезьяны (French, Hernandez-Peon, Livingston, 1955); б — кора мозга кошки (Borenstein, Baser, 1960) с указанием активных точек, стимуляция которых вызывает фокальные потенциалы в РФ среднего мозга (величина точек указывает интенсивность вызываемой реакции); в — кора мозга обезьяны (Segundo, Naquet, Buser, 1955) с указанием областей (черные кружочки), стимуляция которых вызывает электроэнцефалографическую реакцию пробуждения; г — кора мозга кошки (Rossi, Brodai, 1956) с указанием областей (штриховка и точки), удаление котор-ых вызывает значительную дегенерацию в РФ среднего мозга. Пунктиром обозначены границы CI и СII.
Если принять условно, что количество кортико-ретикулярных волокон, отходящих от сомато-сенсорной коры и от моторной коры, одинаково, то и в этом случае на уровне ствола мозга при удалении моторной коры исследователь увидит значительно больше дегенерировавших волокон, чем при удалении сомато-сенсорной коры, за счет того, что к кортико-ретикулярным волокнам моторной коры присоединяется большое число волокон пирамидного тракта.
Следовательно, уже на подступах к ретикулярной формации видно явное преобладание числа дегенерировавших волокон, берущих начало в моторной коре. Такая картина очень легко может убедить пристрастного исследователя признать преобладание кортико-ретикулярных связей моторной коры, тем более что не всегда удается установить точно, заканчиваются ли все обнаруженные волокна в ретикулярной формации. Подчеркивая это обстоятельство, мы не стремимся оспаривать вполне вероятную возможность, что моторная кора действительно имеет более тесные связи с ретикулярной формацией, чем сомато-сенсорная (а также другие специфические проекционные области коры: зрительная, слуховая). Мы стремимся лишь отметить, что связи моторной коры с ретикулярной формацией обеспечиваются кортико-ретикулярными волокнами и, возможно, коллатералями пирамидного тракта, а связи сомато-сенсорной коры — главным образом кортико-ретикулярными волокнами. Не исключена возможность, что в одноименной системе связей сомато-сенсорная кора мало в чем уступает моторной.