ИЗБИРАТЕЛЬНЫЙ ХАРАКТЕР КОРТИКОФУГАЛЬНЫХ ВЛИЯНИЙ II СОМАТО-СЕНСОРНОЙ ОБЛАСТИ КОРЫ
В предыдущей главе достаточно подробно представлены результаты исследований, которые продемонстрировали исключительное значение II сомато-сенсорной области коры в ответных реакциях РФ и СМ на соматические афферентные сигналы. Главный итог этих исследований — доказательство того, что блокада специфической проекции данной афферентной системы в области CII путем ее локального охлаждения приводит к значительному снижению амплитуды фокальных потенциалов или прекращению активности одиночных нейронов, вызываемых стимуляцией той же афферентной системы. В связи с этим сразу же встал вопрос о том, как будут реагировать эти нейроны на другие афферентные стимулы.
Исследование этих вопросов осуществлялось в таких же экспериментах при сочетании соматических и зрительных стимулов. Основное внимание уделяли II сомато-сенсорной области коры, так как влияние I сомато-сенсорной области в исследуемых реакциях не проявлялось.
Сначала определяли фокус максимальной активности одного из соматических нервов в CII для последующего охлаждения этой проекции. Затем обнаруживали активный пункт для отведения фокальных потенциалов или экстраклеточной активности одиночных нейронов в РФ среднего мозга или в ядрах таламуса (СМ или ядро R), вызываемых соматическими и зрительными стимулами. Чтобы обеспечить равномерную освещенность сетчатки при фотостимуляции, за 30—40 мин до начала регистрации потенциалов в оба глаза животного закапывали 1% раствор атропина. Для контроля в зрительной коре отводили потенциалы, вызванные фотостимуляцией, до, во время и после охлаждения области CII. Как и следовало ожидать, локальное охлаждение CII в течение 5—10 мин не вызывало каких-либо изменений в конфигурации вызванных потенциалов зрительной коры. Общий план экспериментов был таким, что позволял изучать реакции одних и тех же ретикулярных (или таламических) нейронов на зрительные и соматические сигналы при охлаждении области CII. На рис. 17 видно, что при охлаждении области CII ответы в РФ уменьшаются по амплитуде только на раздражение соматического нерва и не меняются в ответ на зрительные стимулы (А. Г. Рабин, 1964а, 1965; Р. А. Дуринян, 1968). Та же закономерность наблюдалась и в реакциях одиночных нейронов РФ среднего мозга (В. Л. Цатуров, Р. А. Дуринян, 1968) и срединного центра таламуса (рис. 18).
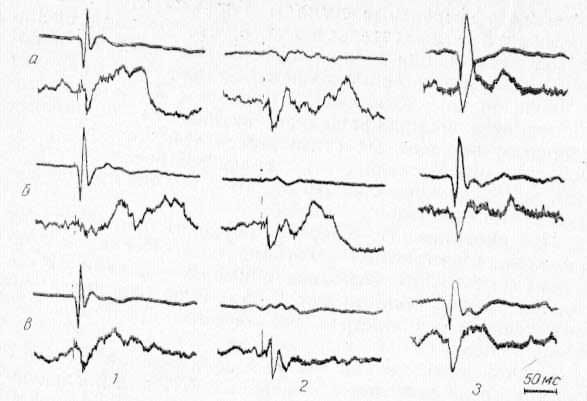
Рис. 17. Избирательность влияния CII области коры при ее охлаждении на потенциалы ретикулярной формации среднего мозга (нижний луч), вызванный при стимуляции нервов контралатеральной стороны (1), нервов ипсилатеральной стороны (2), вспышками света (3). Верхний луч — потенциалы в соответствующих проекционных областях коры (соматической и зрительной). а — до охлаждения; б — во время охлаждения; в — при отогревании
Сходные данные получили Borenstein, Buser (1960) Baser, Bruner, Sindberg (1963), Buser, Richard, Lescop (1969), Richard, Angyan, Buser (1972), хотя, как мы уже указывали, эти авторы не делали попытки дифференцировать влияние первых и вторых зрительных сенсорных областей коры. Они установили, что охлаждение зрительной коры вызывает подавление амплитуды фокальных «зрительных потенциалов» в СМ, в то время как «слуховые и соматические потенциалы» не меняются. То же самое имело место при отведении экстраклеточной активности одиночных нейронов РФ при охлаждении сенсомоторной коры (В. Г. Зилов, 1970, 1972; Buser, Richard, Lescop, 1969) или ее стрихиизации (Meulders, Colle, 1966). Упомянутые исследования дают право утверждать, что обнаруженные нами закономерности в сомато-сенсорной системе могут быть распространены и на кортикофугальные механизмы зрительной и слуховой сенсорных систем.
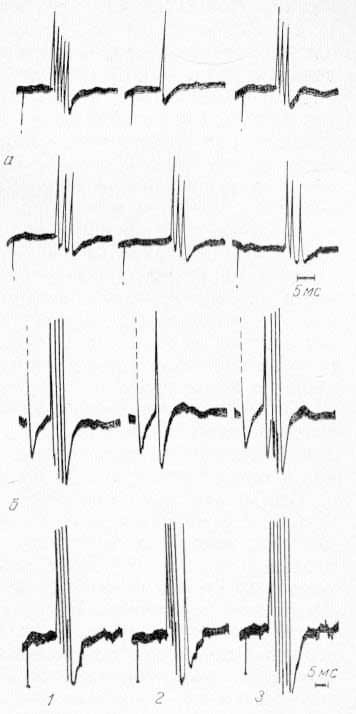
Рис. 18. Избирательность влияния CII области коры на экстраклеточную активность одиночных нейронов СМ таламуса (а) и ретикулярной формации среднего мозга (б), вызываемую соматическими (верхняя запись) и зрительными (нижняя запись) стимулами. 1 — до охлаждения; 2 — во время охлаждения; 3 — после отогревания коры в области CII.
Важно отметить, что эти исследования раскрывают избирательный характер кортикофугальных влияний на нейроны ретикулярной формации и неспецифических ядер таламуса. Каждая сенсорная система своими кортикофугальными сигналами управляет ответными реакциями ретикулярных структур только на сигналы собственной сенсорной модальности, причем механизм этот представлен во вторых сенсорных областях коры. В отношении сомато-сенсорной системы это утверждение основывается на результатах собственных исследований, которые полностью подтверждены недавно (Lindsley, Ranf, Barton; 1972; Lindsley, Rant, Sherwood, Preston, 1972), Для слуховых и зрительных систем основанием для такого заключения (помимо общих итогов исследования Buser и сотр.) может служить обстоятельная работа Ascher (1965, 1966). Автор изучал механизмы так называемой реакции вздрагивания (startle reaction) у кошки под хлоралозовым наркозом в ответ на зрительные, слуховые и соматические стимулы. Анализируя изменения фокальных потенциалов в ретикулярной формации среднего мозга при локальных воздействиях на корковые проекционные области, автор отметил, что нс все участки зрительной и слуховой коры оказывают влияние на ретикулярные ответы. Наиболее эффективными оказались вторые сенсорные области зрительной и слуховой коры. При воздействиях на первые области эти влияния не были выражены. В сомато-сенсорной коре такого анализа автор не проводил и область CII вообще не исследовал.