В неспецифической системе таламуса одной из важнейших структур наряду с СМ является ретикулярное ядро (R), морфофизиология которого была представлена ранее. Это единственное таламическое ядро, которое посылает волокна во все области коры и рассматривается как основной коллектор для выхода на кору всей неспецифической системы. Это ядро получает информацию из многих структур мозга (РФ, гипоталамуса, субталамуса, ядер стрио-паллидарного комплекса и т. д.), в том числе и от специфических передаточных ядер, так как проходящие через ядро R таламо-кортикальные аксоны отдают свои коллатерали на клетки этого ядра. На те же клетки поступает информация и из коры, так как проходящие через ядро R корково-таламические аксоны также отдают свои коллатерали на эти клетки. Тесная связь ядра R с интраламинарными ядрами таламуса, в частности с СМ, и конвергенция на нейронах этого ядра специфических таламо-кортикальных и кортико-таламических путей дают основание полагать, что в ядре R так называемая неспецифическая информация встречается с двумя потоками специфических сигналов: таламическим и корковым. В отличие от срединного центра ядро R получает кортикофугальные волокна от всей проекционной коры и этим напоминает больше ретикулярную формацию ствола мозга. Как структура, расположенная ближе всего на пути кортикофугальной системы, ретикулярное ядро таламуса представлялось важным и интересным объектом исследования.
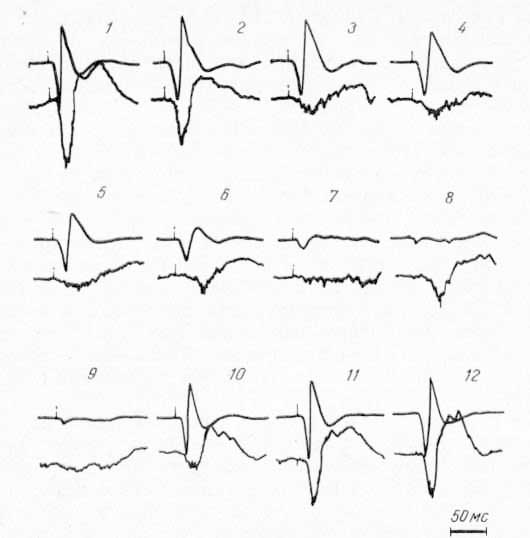
Рис. 21. Изменение потенциалов ядра R, таламуса (нижний луч), вызванных стимуляцией соматических нервов, при охлаждении CII области коры. Верхний луч — запись потенциалов CI. 1 — до охлаждения; 2—8 — во время охлаждения; 9—12 — при отогревании.
При попеременном охлаждении I и II сомато-сенсорных областей коры, как и в случае с другими структурами, наблюдалась прямая зависимость амплитуды ответов ядра R таламуса от ответов II сомато-сенсорной области. При охлаждении СII, в то время как первичный ответ в области CI еще совершенно не изменен по амплитуде, в ядре R амплитуда ответа уже резко подавлена (рис. 21, 3, 4). При дальнейшем охлаждении ответ в ядре R исчезает почти полностью (рис. 21, 7, 9). При отогревании, когда корковый потенциал в CI уже восстановлен (рис. 21, 10), ответ в ядре R только начинает появляться. Следует подчеркнуть, что в ядре R при охлаждении области CII имеет место практически полное подавление вызванных ответов. В других структурах (в РФ среднего мозга, в СМ) фокальный потенциал полностью не исчезал при охлаждении CII; полное подавление имело место дли активности одиночных нейронов.
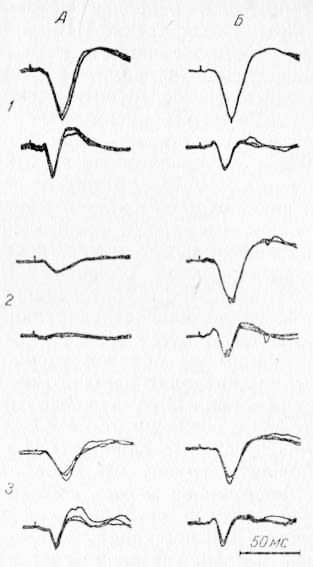
Рис. 22. Избирательность влияния CII области коры при ее охлаждении на потенциалы ретикулярной формации среднего мозга (верхний луч) и ядра R таламуса (нижний луч), вызванные стимуляцией контралатеральных (А) и ипсилатеральных (Б) нервов. 1—до охлаждения; 2 — при охлаждении; 3 — после отогревания.
В РФ среднего мозга, как уже отмечалось, кортикофугальный эффект реализуется полностью лишь примерно на 1/3 исследованных нейронов. Несколько меньшее количество нейронов реагирует снижением ответной активности, но не полным подавлением. Половина ретикулярных нейронов вообще не реагировала на охлаждение CII. Этим, по-видимому, объясняется тот факт, почему фокальный потенциал, вызванный стимуляцией нерва, не подавляется полностью в РФ при охлаждении CII, в то время как реакция отдельных нейронов может подавляться полностью. В СМ эти закономерности специально не проверялись, однако и там встречались нейроны, которые либо не реагировали вовсе на охлаждение СII, либо реагировали частичным снижением активности, хотя по сравнению с РФ общее впечатление таково, что кортикофугальный эффект в СМ представлен более интенсивно, т. е. подавляется активность большего числа элементов. На рис. 22 для сравнения представлены записи потенциалов ядра R таламуса и РФ среднего мозга, откуда видно, что при охлаждении CII в РФ амплитуда фокального потенциала частично сохраняется, а в ядре R подавлена полностью. Однако и в ядре R, как и в РФ или СМ, при стимуляции ипсилатерального нерва, несмотря на охлаждение контралатеральной области СII, отводится нормальный потенциал. Этот факт свидетельствует о пространственной избирательности кортикофугальных эффектов и на ядро R таламуса.
Каким же образом можно объяснить полное подавление контралатерального ответа в ядре R таламуса при охлаждении II сомато-сенсорной области коры в отличие от РФ и особенно от СМ? Следует ли полагать, что кортикофугальные влияния CII передаются на значительно большую группу нейронов ядра R, которые участвуют в формировании ответной активности на соматические афферентные сигналы? Такая возможность не исключается. Ретикулярное ядро таламуса, стоящее «на выходе» неспецифической системы, по-видимому, требует более мощного кортикофугального контроля, чем другие исследованные структуры. Можно напомнить, что на нейроны этого ядра конвергируют одновременно коллатерали аксонов соматического передаточного ядра (VP), посредством которых в ядро могут поступать копии сигналов, передаваемых из VP в CI и CII области коры, и коллатерали кортикофугальных волокон, идущие в CM и другие интраламинарные ядра таламуса, главным образом из области CII. Этим, по-видимому, и объясняется, почему ранний ответ в ядре R, вызванный стимуляцией периферических нервов, имеет некоторое сходство с потенциалами VP и отличается от фокальных потенциалов СМ более коротким латентным периодом и меньшей длительностью фаз. Не исключена также возможность, что в ядре R функциональная организация механизма кортикофугального контроля несколько отличается от организации этого механизма в СМ и РФ дополнительными возможностями для модуляции передаваемых афферентных сигналов. Чтобы обсуждать этот вопрос, следует рассмотреть сначала хотя бы в общих чертах тот механизм, который, согласно нашему представлению, может лежать в основе реализации кортикофугальных эффектов на нейронах неспецифических ретикулярных структур мозга. Для краткости, будем именовать эти нейроны, как и раньше, ретикулярными нейронами в тех случаях, когда не будет необходимости специально подчеркивать, о какой структуре идет речь.